細胞質遺傳特別對母系血統的研究
緒 言
人、家畜、子代係由父及母所生的,通常係有在其「親」到育成時期為止。假使以被生的子去推定到離乳時為止的時期,如以離乳時對子的體重去考慮時,父與母任何時候是否皆有較大的給與影響子代的離乳時體重?如被認為係母對子的授乳狀況,母的影響是比父的影響大。是因母對子給與母乳而育成的。乳牛的情形,在剛出生時由母的初乳給與子,母的影響可認為比父的影響大。但是不給與初乳時到底如何?包含初乳、乳由母給與、多少量的乳供飲用,在子是有環境的問題,乳不由母給與環境下,依然母的影響方面仍然被考慮比父大。此係哺乳動物的情形,也就是子在出生前胎兒是在子宮中發育。
以上係子的離乳時體重受父母的影響有取自環境的情形,那麼環境的要因與遺傳的對比到底如何?在一般的子係由父與母各得一半的遺傳因子。是否應該如此?在確實的,如限於性細胞的核內遺傳子 ( nuclear genes ) 一般是如此。但是遺傳因子及其構成的 DNA 不僅在核內,核也有包圍細胞質 ( cytoplasma )。在細胞質內有種種微小器官 ( organelle,例如葉綠體 chloroplast ),其中粒線體 ( mitochondria ) 是很大,可知存在著很久遺傳因子。細胞質在母的性細胞有卵子,在公的性細胞有精子。子係由父及母得到均等的核內遺傳子,由母的細胞質或粒線體而來的遺傳子有關係時,在遺傳上母的影響比父的影響大。
上面的例係以離乳時體重去推定,子的任何的形質 ( character, trait 例如身長 ),能力 ( performance 例如對屠體的肌肉量 ),以及遺傳與環境的二因素之決定是一般的考慮。尤其個體的遺傳效果 ( genetic effect ) 與環境的效果 ( environmental effect ) 由個體的表型值 ( phenotypic value:測定值 ) 決定之,因個體間被看到不同而引起變異 ( Variation )。
到此尚有當初的質問變動,子對遺傳、環境也受到父及母很多的影響而得到答案。在子尤其環境的要因對泌乳的影響,係在離乳後慢慢的消失,對胎內的影響一般也沒有看到延長,遺傳子的影響之形質,由母而來的遺傳子比由父而來的多,一般,可考慮母對此的影響比父的影響大。由此上所述再敘述,如下的說明。
子代的表現型 ( 值 ) 之決定有公與母。子係由父與母接受均等的核內遺傳因子,由母重新的接受細胞質而來的遺傳子。在環境的係子再包含接受母的泌乳性之能力 ( mothering ability,母的能力,例如與育成有關的行為的影響 )。
本文所使用的各種述語。如母性效果 ( maternal effect ),母的影響 ( materal influence ),細胞質遺傳 ( cytoplasmic inheritance ) 之類。除此之外的定義,例如母的影響 ( influence ),在子的表現型值成為母的效果 ( effect ),可由子的表現得到說明。所謂定義,定義隨人或群而有所不同,常常在其間有所混亂,讀者須理解正確的問題本質,有些定義以靈活回轉的方式為宜。在一般因短的術語之亂用而招致無用的混亂。本文係以附合細胞質尤其粒線體之遺傳研究成為焦點敘述如下。
粒線體 ( mitochondria ) DNA ( Mt DNA ) 的性質
哺乳動物的粒線體幾乎係生產細胞活動所需全部的熱能,被稱呼是好細胞的熱能庫 ( powerhouse )。在此情形粒線體係對熱能源負有重要的任務,因此粒線體的遺傳子或遺傳子群 ( 遺傳子型 ) 之變異,而產生個體的能力之間的差異是否沒有關係的質問。若兩者之間任何的關係被看時,是否可單就基於粒線體的遺傳之變異進行選拔?又在選拔基率有關的工作能力資料之組合,是否能夠用於促進家畜的遺傳改良?DNA 以旋轉技術的急速進步的今日,mt DNA 的遺傳子型判定僅在研究室便可進行。
哺乳動物的粒線體遺傳子像 genome ( 全體的遺傳子像 ) 係二重螺旋 DNA 構造有 16.5 kilobase 單位的長度。mt DNA nucleotid 配列,人、牛、小白鼠係相同的。所謂 D loop 領域 ( D-loop region ) 被稱呼的領域,人與牛之間的主要 DNA 的長度及數量不相同。其 mt DNA 領域在家畜進化之間沒有變化,在機能的被考慮是很重要的。人也是 mt DNA 的突然變異而引起退行性疾病及老化,粒線體異狀成為疾病的分子生物學的基礎問題。人有以含動物體之性細胞 ( 如精子 ) 之外的體細胞 ( 如肝細胞 )。在體細胞引起很多 mt DNA 突然變異,而促進老化及疾病。肝的粗線體活動在老年時降低,與老化過程有關係。小白鼠系統間一生的生產能力有顯著的差異性,大概可能與粒線體的不同有關。
在家畜對細胞質的遺傳子像 ( genome ) 係以 mt DNA 為主體而構成的。不論公或母,粒線體以由母而遺傳之 ( 母性遺傳 maternal inheritance )。牛及小白鼠的卵子大概存在有 10 萬的粒線體,而精子只有約 75 程度的粒線體。因此父性與母性的 mt DNA 量之比大約 1:1000。故精子與卵子結合之結合體 ( zygote ) 之製造時,精子的 mt DNA 有顯著的少。因此精子的粒線體於受精後不久接合體的細胞質內被認為似退行。也因此由卵子接合體的粒線體成為構成的全體。以上 mt DNA 在原則上是由母而來的,由父而來的幾乎沒有。但是 Gyllenstein 等 ( 1991 ) 報告稱母小白鼠以公小白鼠側系統小白鼠,經過 26 代回交 ( Backcrose ) 父性 mt DNA 量在實驗是增加的,父性 mt DNA 量傳給子。
粒線體的遺傳子係與核內的遺傳子不同,因沒有組換 ( recombianation )。mt DNA 係在細胞分裂之際獨自分離 ( segregate ) 復生 ( replicate ) 之。在家畜對細胞內粒線體的數量,係種 ( species ) 泌乳的時期,在組織細胞的變異時期表示有顯著的變異。例如以牛胎兒肌肉細胞去比較,在卵胞細胞 ( follicular ocytes ) 有近 100 多倍的 mt DNA 存在。此情形在牛的卵發生期 mt DNA 也增大 100 倍。在腦、肝、乳腺之類的新陳代謝須要高熱能的組織,故有多量的粒線體。粒線體其在細胞或個體內有很強的均質性 ( homoplasmic, homogeneous )。不均性粒線體很稀少的原因可由此瞭解。
mt DNA 係由母傳給子。此時突然變異 ( mutation ) 也有引起部份的變化,在一般由母傳給子同樣也傳給 mt DNA。一方面,對於核遺傳子,子係由父及母接受遺傳子而繼續,所謂遺傳子的分離 ( segregation ) 及組換 ( recombination ) 之引起,親與子係完全持同一的遺傳子型。但是親子鑑別使用 DNA 試驗時,以調查 mt DNA 為宜。何故?在原則上子係持有與母同一 mt DNA。過去,有報告樺太離別後母與女兒經過的 50 年後再相會,有使用 DNA 試驗的消息。大概是以粒線體的 DNA 供調查?mt DNA 其情形在實際方面也是相當有趣的 DNA。
更一進步的,又觸及粒線體遺傳子與母性效果 ( maternal effect ) 的問題。例如母的泌乳性之類的表型能力如前述的,母的遺傳與環境效果之總和表現被考慮。在遺傳要因是在核遺傳子之外,粒線體、遺傳子是當然的被考慮。其是因全部的遺傳子總合體被置在母的環境下之泌乳被發現泌乳是包含形成子的哺乳時環境。在泌乳時乳腺是必要多量的熱能,對粒線體的介入是有很大的意義。在子的成長形質對母性效果的情形時,子成長有關遺傳子與母的泌乳有關的遺傳子是有關係,又更進一步的,如加上母與子的環境要因時則稍微的低。所謂母性效果是在給與母與子的環境問題,所謂母系血統是在於細胞質,是能力的遺傳效果問題。
母系血統
1.定義
母系血統 ( maternal lineage ) 在一般的,以追尋血統 ( pedigree ) 表到原集團 ( foundation population ) 為止,追溯全部的母系而被樹立。在被登錄一頭的原集團母而來的全部後代 ( 彼等係以母系血統個體稱呼 ) 係母系血統 ( 也就是母性血統 ),母系統 ( maternal line ) 或細胞質品系 ( cytoplasmic line ) 是屬於個體而被整理。總之是原集團對於一頭登錄的母係此等的母系血統群全部的個體之共同祖先。在母系血統群與能力 ( 例如泌乳量 ) 之間是否被看到相關關係?在調查統計分析之際,以使用母系血統群為固定要因 ( fixed factor )。
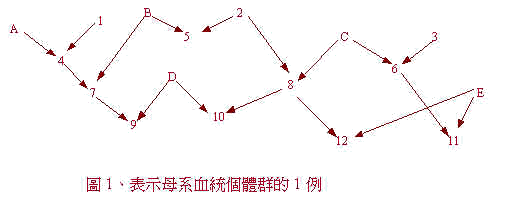
在圖 1 係表示成立母系血統個體群的情形。其血統圖表 ( chart ),字母表示公的,數字表示母的,若假設基礎母 1、2 及 3 無血緣 ( 正確的粒線體、遺傳子型 ) 關係時,母系血統個體群是有 3 個。例如對母的限於形質 ( 泌乳能力等 ) 的分析,母系血統個體群 1 係母 1、4、7 與 9,個體群 2 係母包含 2、5、8、10 及 12。若資料由數代收集時,在統計分析型式係考慮世代效果,母的出生年或世代效果非考慮不可。若記錄公的及母的能力 ( 例如成體重 ) 時,( 總之非限性形質 non sex - limited trait 時 ) 兩性的能力以母系血統個體群去考慮行統計分析。但是此時性別要因係以模型進入,對性別不同能力的除去。血統圖表如能作出來,母系血統個體群使用電腦便容易設定。C.Y.Lin 等,預定以 Fortran 電腦預定表作小白鼠 52 世代有關的繁殖壽命資料去設定母系血統個體群。
在談若干觸及與 maternal lineage 有關連的 paternal lineage ( 父系血統 )。日本及西歐諸國,在一般的女性的結婚冠上夫的姓名,結婚前的姓係少女的姓名 ( maiden name ) 不被置入。此種習慣是相當古老的。因此此種情形在農業種下各種根。煙草的收獲能夠高,係受到種的良否所左右。同樣的有良好的子及不良的子之分別,夫的配種 ( 過談問題 ) 之決定被考慮。因此在過去日本有借腹生產之發生,女的出嫁生子是最大的義務。此種表現是蔑視女性,在缺乏繁殖及遺傳知識時代,此種背景產生女性的結婚須冠夫姓的社會狀況 ( J.W.Leth )。無論如何對於生物學在 maternal lineage 係意味著與 DNA 有關係,在 paternal lineage 除追尋男性方面的血統外,其他是沒有意義的。
2.有關細胞質遺傳效果的文獻
在離乳時前成長與乳量對細胞血統群間看到有顯著的差異。但是 Tess and Rabinson ( 1989 ) 稱肉牛不能確認其效果。Bell 等稱乳牛的乳量與乳脂量等的變異之中,1.8 ~ 3.5% 是由細胞質的變異而來的。Hwzinga 等 ( 1986 ) 稱乳量等的變異之 10% 係細胞質所決定。母系血統效果乳量是 5.2%,乳脂量是 4.1%,因此乳脂量也達 10.5%。mt DNA 的多型性 ( polymorphism ) 對乳量,乳脂持有極重要的意義。母系血統效果對乳脂率與乳生產熱能量 ( milk energy concentration ) 是很重要的。此等的研究係細胞質性的母系血統效果,表示乳牛係有潛在的有用的變異。但是一方面 Kennedy ( 1986 ) 與 Reid and Van Vleck ( 1987 ) 稱乳量等的形質是很重要的細胞質遺傳要因。Ron 等 ( 1993 ) 稱 israel holstein 324 母系血統群之中最高與最低乳量之 4 母系血統群比較其平均值,2 群之間對 D-loop ( D–輪 ) 領域的多型變異是沒有不同。
家畜包含哺乳動物在調查其細胞質效果的存否,相互雜交母 ( reciprocal cross females ) 間之能力以同期比較 ( contemporary comparison ) 方法是可以使用的。但是相互雜交公的之間之同期比較,伴性 ( sex-linked ) 遺傳與細胞質效果混合 ( confounding ) 被認為是不可以的。相反的,在家禽是由母系的決定性。F1雜交係雜種強勢 ( heterosis ) 的評價,F1的平均以兩親系統的平均與比較去進行,在其推定值係混合母方的細胞質效果 ( 參照母性效果的說明 )。
基於哺乳動物的母系的記錄 fullsib ( 全兄弟 ) 分散分析時,若父的分散 ( sire variance ) 比母的分散 ( dam variance ) 大時,性聯遺傳 ( sex linkage ) 表示有其效果。分散分析 dam variance 比 sire variance 大時其不同差是細胞質效果,那麼在性聯遺傳效果,遺傳子的優性 ( dominace ),必須考慮到包含遺傳子相互作用 ( epistasis )。使用過去的交配方法、性聯遺傳、優性甚至要切離相互作用效果,以細胞質效果,推定最困難的。
3.對於母系血統效果分析之假定
在資料分析時,全部的登錄基礎母假定由不同的細胞質源而來。Brown 等 ( 1988 ) 等報告由 mt DNA 的分子生物學的分析,在血統依其方法係真的細胞質血統 ( cytoplasmic lineage ),據報告也許尚未充分也未知。有登錄基礎母由相同的母祖先而來大概是有的。此情形時,單一的共同之母系先祖後代不同的幾個母系血統群被分開,成為很多不必要的母系血統群提供試驗。何種程度的母系血統群被誤認,以統計結果要預知其缺失是困難的。
再說母系血統群間的能力差以試驗統計分析,認為係同一母系血統群內的粒線體遺傳子引起突然變異而在各世代遺傳,同一母系血統群內的 mt DNA 係均一 ( homogeneous ) 的,群間係不均一 ( heterogeneous ) 的而被考慮之。因此突然變異的引起,頻度 mt DNA 比核 DNA 高。mt DNA 配列的變異係存在於母系血統個體群內。若 mt DNA 突然引起變異時,母系血統個體群 mt DNA 的均一認定被崩潰。故 mt DNA 突然變異,在能力以其為重大效果時,突然變異效果對修正進行有限,突然變異效果與真的母系血統個體群間之差形成混合。mt DNA 突然變異對乳牛是很重要的問題。乳牛的母系血統個體群在全國的遺傳之評價時是普通的,血統 ( 血緣關係圖表 ) 是依據此圖而設定的。血緣個體的線如很長時,基於突然變異與血緣分類失敗的機會很高,故很多母系血統個體群是用不同的分類。
母牛大部份的 mt DNA 突然變異係 D-loop ( 非符號化 ( coding ) 領域 ) 引起的。若有突然變異是母的 mt DNA 引起的,資料統計處理時,其母與後代個體係被別的 mt DNA 引起的,資料統計處理時,其母與後代個體係被別的母系血統個體群所吸收。對於此問題的理論係此情形的 sublines 之間的能力不同,到底如何有須再加以試驗研究。若 sublines 之間沒有差異時,此等 sublines 係單一母系血統個體群之被正當化。突然變異與錯誤的血統圖表對母系血統個體群之分類應避免其過失,使用血統圖表以併用 mt DNA 為宜。
Kennedy ( 1986 ) 的電腦使用的模擬試驗有所表示,選拔與 drift ( drift,在小個體群內對遺傳子頻度的世代間彷徨變異 ),係細胞質性血統效果 ( cytoplasmic lineage effects ) 成為正的指定妨害。但是母系血統個體群間的選拔資料有行適當的統計修正的必要。所以適度的大小之母系血統個體群之使用,drift 有迴避的必要。在基於定義上,母系血統個體群內設定是同一粒線體時,此粒線體的 drift 係本來的問題以外者。
4.粒線體與核遺傳子間的相互作用
在粒線體對熱能生產徑路,係粒線體及核的遺傳子因被控制,核的遺傳子與粒線體的遺傳子之間有互相的關係。但是細胞質的家畜能力對其效果的研究,幾乎的情形是在兩者假定沒有相互作用。其原因兩者效果的固定尚屬困難。其次是對此情形的相互作用的性質在家畜必須瞭解才可。
母的核遺傳子與粒線體的 genome 係共同的被遺傳給子,核遺傳子,粒線體的遺傳子其相互作用的效果是混合的,過去的交配系統之下分離的推定是困難的。但最近核移植技術的被開發、DNA 型態分類成為可能、相互作用的效果也成為能測定出來。簡單的說明以二個系統 A 與 B 去設定。A 系統的個體之核內遺傳子是有 AA 的遺傳子型,同樣的在 B 系統是 BB。核移植以 A 系統與 B 系統進行,AA:mtA、AA:mtB、BB:mtA、BB:mtB 之 4 個的核,粒線體能夠組合遺傳子型。AA:mtA 與 BB:mtB 的遺傳子型係不能核移植,系統內交配能夠得到。無論如何有如下 3 個很趣味能夠作比較。
1.AA:mtA 與 AA:mtB 的平均及 BB:mtA 與 BB:mtB 的平均比較,系統 A 與 B 之間的核遺傳子型值的差能夠推斷。
2.AA:mtA 及 BB:mtA 之平均與 AA:mtB 及 BB:mtB 的平均比較,A 與 B 系統間的粒線體遺傳子型值的差能夠推斷。
3.AA:mtA 與 BB:mtB 之平均與 AA:mtB 及 BB:mtA 的平均比較,核與粒線體間的相互作用效果能夠推斷。
AA:mtA 與 BB:mtB 的比較,係以 A 與 B 系統間之核,粒線體遺傳子型的總合效果去推斷須注意。此係由過去的交配下系統差調查時的典型比較。以此情形比較時,在統計原文 ( text ) 的比較 ( contrast ) 之文章內認為有連帶關係。
相互雜交母 ( reciprocal cross females ) 之間的能力被認為有顯著差異時,係普通細胞質的差是可解釋明白的,又假定核、粒線體沒有相互作用。相互雜交間公的比較,對細胞質的差沒有給與正確的評價。為何如此,因在核、粒線體假定沒有相互作用之外,細胞質效果是由性染色體混合而來的。使用核移植法,相互雜交公的核以相同母系血統個體的卵子移植也可以。因此以此 2 組的相互雜交公係持有相同細胞質背景,在欠缺細胞質要因的效果比較,是能夠推定係沒有關係的性染色體效果。
細胞質遺傳之遺傳育種學的意義
若以 mt DNA 多型是家畜的能力之差的重要任務為結果時,對供作為家畜的遺傳改良是有用的。
1.為著後代指定的公及母 ( bull dam ) 之選拔及為著胚移植、cloning ( 在組織培養以遺傳的作出與親代相同的個體 )、分割的母系選拔時,在核遺傳子型加入母的粒線體遺傳子型也有考慮的必要。任何的卵子,其母全部的後代由 mt DNA 遺傳。
2.對於公與母的粒線體遺傳子型有關預備知識,對公評價 ( sire evaluation ) 的精度提高是沒有用的。何故?目前在日本的全國的遺傳評價系統,對公的育種價之正確評價全部已得到。種牛的粒線體遺傳子型已瞭解的是粒線體由母傳給子,在遺傳的改良沒有任何的利益。
3.喜好粒線體遺傳子型頻度的增加,由優良母牛的細胞質而來的粒線體在 ( 受精 ) 卵能夠微量的注入。
4.二元或為著多元交配製造優良的 mt DNA,雞及豬行二元或多元交配時,朝向市場的家畜生產,在實際上是很有趣味的。
5.遺傳的助變數 ( parameter ) ( 例如遺傳率及遺傳相關 ) 之推定,總是有母性遺傳效果。因此要考慮細胞質效果時須以母系血統效果為目標是屬於很重要的。細胞質遺傳係母方血緣動物模彷的增加,母方一半兄弟是比父方一半兄弟更相似。其結果,女兒,母回歸或母方一半兄弟的分析,係兒子,父回歸,女兒,父回歸及父一半兄弟分析比較,可得到高遺傳率與遺傳相關的推定值。
6.高率的 mt DNA 突變變異,一卵性雙生子 mt DNA 不同也未知,在一般被想定是同一遺傳材料所造成的也未知。若 mt DNA 的突然變異顯著影響能力時,持有同一遺傳材料時考慮由行一卵性雙生子的實驗結果得到解釋。mt DNA 係由母傳給子,母方的性細胞品系 ( female germ line ) 之粒線體突然變異係全部由母方傳給子,因此持著重大的遺傳結果而加以附記之。此在對照的,體細胞 ( somatic cells ) 或哺乳動物公性細胞品系之粒線體突然變異,係其子因得不到遺傳,而沒有任何的問題。
7.有文獻對此的報告,若細胞質的遺傳重要,種牛的育種價推定值 ( estimated transmitting ability, ETA ) 係限於在正當時不被解釋,成為偏見 ( bias )。此種情形是種牛的 ETA 被推定以其母方血統個體的能力資料為基礎,在細胞質對 ETA 的一部份是因不遺傳給其兒子及女兒。被行任意的交配時,種牛不分母方係優秀的母牛或係不良的母牛皆被交配。交配母牛的粒線體遺傳子型在一般交配前須預先瞭解,交配係一般的有關細胞質遺傳是非常的無規則的。
摘 要
很多的研究報告認為細胞質的遺傳對家畜的能力變異有關。細胞質遺傳是家畜生產改良以遺傳的手段給與的另一個視角。本文係以家畜育種對細胞質遺傳的意義之敘述。細胞質係與核 genome 同程度,可知是對能力有重大的影響,粒線體情報能夠使用由過去的選拔育種為補助方法。為著提高喜好粒線體的頻度,卵或胚細胞質的微量注入 ( microinjection ) 也可以使用。細胞質 genome 效果的研究,係細胞質效果是普通性聯遺傳 ( sex linkage ) 與核,細胞質遺傳子相互作用效果在與其混合時受到妨害。目前的情形,僅是家畜的相互作用之被理解為止而已,尚期待今後的繼續研究。核移植等的方法之使用係今後核遺傳子型,粒線體遺傳子型其相互作用的效果能夠個別的推定。以母系血統個體群間的能力差異與細胞質遺傳的關係去調查時,以對粒線體的突然變異與登錄基礎動物的血統圖表之正確度去考慮,並行深切的注意之。由粒線體情報得到的經濟利益是 mt DNA 的判定,分類所需費用多少「錢」非考慮不可。
( 康天旗 )
中國畜牧雜誌第30卷(98)第 6 期 ( 9 ~ 18 )