學歷:國立台灣大學畜牧系畢業
現任:省立宜蘭農工職校畜牧獸醫科教師
五、氨基酸平衡
(1)良好的氨基酸平衡
Mitchell (1946) 認為蛋白質的生物價與氨基酸含量的相關性很高,且蛋白質的營養價是由其所含的氨基酸而來的。此後Scott (1969) 證明雛雞,老鼠等即使飼料中不含蛋白質,但若含有適當量之氨基酸時即能得到正常的成長,且氨基酸代替蛋白質的營養價。
欲氨基酸之混合物呈現最高的營養價,必需其所含之氨基酸的量的相對的比率適當,此種相對的量的比率稱為氨基酸平衡 (amino acid balance)。氨基酸的要求量是以飼料中的量表示,而氨基酸平衡則多以每氮素16 g,即每粗蛋白質100 g的量表示。
Mitchell認為營養價最高者為全卵蛋白質,將其生物價當做100,此並非表示全卵蛋白質的氨基酸平衡與必須氨基酸要求量的相對量一定相等。
Williams等 (1956) 最先研究最適當之氨基酸平衡。其基本的想法是與在組織或血液等動物體內合成的蛋白質中所含的必需氨基酸的比率相同的氨基酸平衡為最適當者。William等進行鼠、豬、雞之屠體分析,並在實驗上得到? 這些動物之屠體的必需氨基酸含量互相很接近, 由增重及屠體分析值來計算的必需氨基酸之要求量的數值與由成長實驗所得到之要求量的數值很一致, 屠體之氨基酸組成與氨基酸要求量的數值很接近等結果,並主張能從屠體分析測定氨基酸要求量。
另外,Mitchell對各種蛋白質的生物價就幼鼠、成鼠、成人測定數值而加以比較,發現即使是同一蛋白質離氨酸含量少者其生物價為
成鼠>幼鼠,成人
甲硫氨酸含量少者為
幼鼠,成人>成鼠
而且,視所合成之蛋白質的氨基酸含量而良好的飼料氨基酸平衡不一樣,變成接近所合成之蛋白質的氨基酸平衡。
此種場合,成長旺盛的幼鼠及被毛少的人以肌肉蛋白質的合成為主,從而離氨酸的要求量相對的變大,離氨酸含量少的小麥麩質 (wheat gluten meal) 的生物價相對的變低。而成鼠方面,被毛成長上的負擔相對的變大,甲硫氨酸+胱氨酸的要求量增大,故甲硫氨酸+胱氨酸含量少的花生粕生物價相對的變低。
Scott根據此種想法,進一步以次式計算形成體蛋白質而蓄積的氨基酸量,並與要求量的數值做對比,算出各個必需氨基酸的效率。
表5-1 全卵蛋白質之氨基酸含量與雞之氨基酸要求量
全卵蛋白質 |
氨基酸要求量.飼料蛋白質中 |
||
雛 |
產卵雞 |
||
arginine |
6.4 % |
5.0 % |
5.0 % |
lysine |
7.2 |
5.0 |
4.2 |
histidine |
2.1 |
2.0 |
2.0 |
methionine |
3.4 |
2.0 |
2.0 |
cystine |
2.4 |
1.5 |
1.6 |
tryptophane |
1.5 |
1.0 |
1.0 |
phenylalanine |
6.3 |
3.5 |
4.6 |
leucine |
9.2 |
7.0 |
7.5 |
isoleucine |
8.0 |
4.0 |
5.0 |
threonine |
4.9 |
3.5 |
3.7 |
valine |
7.3 |
4.3 |
4.3 |
氨基酸的蓄積量=增重量×0.18 (a)×(屠體蛋白質中的氨基酸含量) + 羽毛
增加量×0.82(b)×(羽毛蛋白質中的氨基酸含量)
〔(a):單位為g,(b):增重量,增加量中之蛋白質g數〕
表5-2 屠體之必需氨基酸含量
屠體蛋白質中 (%) |
||||
豬 |
雛 |
平 均 |
||
arginine |
5.89 |
7.12 |
6.71 |
6.57 |
histidine |
2.16 |
2.65 |
1.96 |
2.27 |
isoleucine |
3.49 |
3.84 |
4.12 |
3.82 |
leucine |
6.46 |
7.14 |
6.63 |
6.84 |
lysine |
7.61 |
8.55 |
7.46 |
7.87 |
methionine |
1.71 |
1.77 |
1.76 |
1.75 |
methionine +cystine |
3.20 |
2.78 |
3.51 |
3.16 |
phenylalanine |
3.69 |
3.77 |
3.95 |
3.80 |
threonine |
3.87 |
3.79 |
4.02 |
3.89 |
tryptophane |
0.76 |
0.74 |
0.77 |
0.75 |
valine |
5.51 |
6.00 |
6.72 |
6.08 |
tyrosine |
2.88 |
2.59 |
2.49 |
2.65 |
表5-3 鼠及人之生物價
生 物 價 |
|||
幼 鼠 |
成 鼠 |
成 人 |
|
卵白蛋白 |
97 |
94 |
91 |
全卵粉 |
87 |
82 |
94 |
牛 肉 |
76 |
69 |
67 |
小麥麩質 |
40 |
65 |
42 |
酪蛋白 |
69 |
51 |
56 |
花生粕 |
54 |
46 |
56 |
表5-4 使用於組織、羽毛、卵之蛋白質合成的氨基酸的比率 (%)
肉 雞 |
產 卵 雞 |
|||
雛雞 |
期別Ⅰ |
期別Ⅱ |
||
甲硫氨酸+胱氨酸 |
68 |
60 |
--- |
--- |
甲硫氨酸 |
--- |
--- |
76 |
79 |
離氨酸 |
73 |
71 |
84 |
84 |
異白氨酸 |
--- |
72 |
82 |
84 |
纈氨酸 |
--- |
85 |
--- |
--- |
必需氨基酸的作用並非全部成為蛋白質的合成材料,例如甲硫氨基酸分解後即成為甲基 (methyl) 供與體,而具有重要的作用,必需氨基酸不一定優先使用於蛋白質的合成,剩餘的部份才被分解。Scott以前述的方法算出各種必需氨基酸使用於蛋白質合成的比率,且其效率的數值為
纈氨酸>離氨酸>甲硫氨酸+胱氨酸
Scott推測效率低的氨基酸在蛋白質合成以外之生理的作用上被使用的比率多。
最適當之氨基酸平衡,係意味其最適合於飼養目的,且與蛋白質合成量的相關性很高,但並非相同。
體蛋白質被分解後,有相當大的部分被再合成。即被分解之體蛋白質內所含的必需氨基酸有相當大的部分被再利用。此種被再利用的比率視氨基酸的種類而異,而且即使是同一種氨基酸亦視由飼料所供給的量而異。此點最能顯示出某一種必需氨基酸在飼料中的含量不足時此氨基酸的飼料內添加效果視氨基酸的種類而異。
色氨酸缺乏的場合,添加色氨酸時少量的添加即見到顯著之氮素蓄積量的增加,但離氨酸缺乏的場合離氨酸的添加效果就比較少。這是因飼料蛋白質的水準使最適當的氨基酸平衡不同,從而第1限制氨基酸不同所致。
此種必需氨基酸之缺乏領域的變化可再由蛋白質的生物價與化學值的關係來看。膠質 (gelatin) 的色氨酸含量為0,因此化學值 (chemical score) 為0,但由氮素出納來看時營養價即使是單獨使用亦不會是0,生物價為25。離氨酸缺乏方面生物價40,其他一般必需氨基酸缺乏為20。含硫氨基酸的缺乏方面接近0,但因含硫氨基酸尚具有其他重要作用,故是否因蛋白質合成所需的材料不足其生物價才是0並不明瞭。
惟一般人都認為必需氨基酸缺乏時蛋白質合成量不會立即與蛋白質無給與同水準。若必需氨基酸不在體內合成時就不會有此種想法。
因此,由生物價的算出法來看時,以無蛋白質飼料給與時的氮素排泄量,即內因性尿氮素量為基準,先算出外因性之尿氮素,然後間接的算出用於蛋白質之合成的飼料由來的氮素量。因內因性尿氮素量從飼料而來之氮素的吸收為0,故考慮到體蛋白質被分解,再利用時之必然的損失,其量比給與飼料蛋白質小,故體蛋白質在被分解再利用時之營養價在蛋白質中最高,其生物價當做100,並以此為標準。
此蛋白質若具有理想的氨基酸平衡時,即使添加各個之必需氨基酸亦無使尿中氮素量更減少的效果。但動物給與無蛋白質飼料時,添加甲硫氨酸可使氮素出納獲得改善,此點在鼠、豬、雞方面已得到證實。
另外,吉田 (1974) 發現老鼠給與無蛋白質飼料時除了甲硫氨酸之外,羥丁氨酸亦有添加效果,且內因性氨基酸之第1限制氨基酸及第2限制氨基酸乃是這兩種。村松 (1983) 對雞進行實驗,亦發現由於甲硫氨酸的添加使無蛋白質飼料給與時的體重減少獲得改善,而且甲硫氨酸與精氨酸組合起來效果更佳。
瞭解最適當之氨基酸平衡的第二種方法是以氨基酸混合物代替飼料蛋白質使用,其中之必需氨基酸分別拿掉一種而做為基礎飼料,並階段性的加入此種氨基酸,劃出所謂 dose response curve,此曲線形成水平點之添加量當做要求量,對各種必需氨基酸以此種方法作出要求量的表,具有此相對比者即是最適當的氨基酸平衡。
此種實驗之基礎飼料亦有使用缺乏特定之氨基酸的蛋白質,例如膠質或小麥麩質者。在判定上動物側之指標幼齡動物為成長,成長的動物為氮素出納。利用此種方法研究鼠、雛雞、產卵雞之最適當的氨基酸平衡,老鼠方面以Rama Rao之氨基酸型態,雛雞方面以Scott之reference diet最有名。
蛋白質與氨基酸混合物是不相同的。氨基酸混合物在消化管內呈現蛋白質之數十~數百倍的滲透壓,且吸收的形態、經路、速度亦與蛋白質的場合不一樣。但氨基酸混合物與蛋白質,若其氨基酸組成相同的話仍然呈現大體相同的營養價,由氨基酸混合物進行的實驗可提供最有力的資料。
(2)氨基酸之相互作用
1. 4種相互作用
在計算飼料之配合比率時,通常是以氨基酸要求量與飼料之氨基酸含量,飼料原料的價格做為基礎數值,但考慮氨基酸之相互作用亦極為重要。氨基酸之相互作用的結果具有廣泛之生理學的,生化學的影響,並可能引起成長的抑制,飼料攝取量的低下,病理學的變化等種種現象,有時還會招致飼養成績的低下。
Harper (1978) 將氨基酸之相互作用分為主要之4群,即(1)缺乏,(2)不平衡,(3)拮抗作用,(4)毒性。這些是引起成長及其他之飼養成績低下時由飼料之構成面來分類者,而非從其作用機構來分類者。而且在實際上亦有幾種作用同時作用的情形。
表5-5 Scott之reference diet
氨基酸混合物 |
玉米澱粉 (corn starch) |
56.15 |
|
L-arginine.HCl |
1.21 |
玉米油 |
15.00 |
L-histidine.HCl.H2O |
0.41 |
混合鹽 |
5.37 |
L-lysine.HCl |
1.19 |
NaHCO3 |
1.00 |
DL-methionine |
0.35 |
纖 維 |
3.00 |
L-cystine |
0.35 |
氯化膽鹼 |
0.20 |
L-phenylalanine |
0.50 |
維他命混合物 |
|
L-tyrosine |
0.45 |
ethoxyquin |
|
L-tryptophane |
0.15 |
蛋白質當量 |
14.5% |
L-isoleucine |
0.60 |
ME |
4,300kcal/kg |
L-leucine |
1.20 |
||
L-valine |
0.82 |
||
L-proline |
0.20 |
||
L-threonin |
0.65 |
||
glycine |
1.20 |
||
L-glutamic acid |
10.00 |
2. 氨基酸之缺乏
一種或數種必需氨基酸其要求量未獲得滿足時稱為缺乏 (Deficient)。實際之飼料方面特定之必需氨基酸的含量不會是0,故應稱為不足才對,但通常都稱為缺乏。最不足的氨基酸稱為第1限制氨基酸,其次不足的氨基酸稱為第2限制氨基酸,而添加這些氨基酸時成績得到改善。
此種場合由氨基酸之添加效果來看時,單是1種氨基酸不足的場合,效果可達到此氨基酸之要求量的線,但有數種氨基酸缺乏時第1限制氨基酸的添加其效果可達到相當於第2限制氨基酸之充足率的量,超過此以上的添加亦難望有效果。由於氨基酸的添加效果受到其他氨基酸量的影響,故看起來比較單純之單一的氨基酸缺乏的場合亦要考慮相互作用。
3. 氨基酸之不平衡
不平衡的場合,係兩種或數種氨基酸之量的比率使由氨基酸之絕對量所期待的成績低下。在實驗上添加第2限制氨基酸時會形成成績低下的現象。此種場合若添加第1限制氨基酸時成績的低下即解消。
表5-7是表示幼豬給與5-6所示之CP 12%的玉米。大豆粕飼料時之日平均增重量者。飼料之第1限制氨基酸為離氨酸 (要求量的71.4%),第2限制氨基酸為色氨酸 (要求量的83%)。色氨酸的添加會使增加重量減少。此種效果稱為不平衡 (inbalance)。
表5-6 幼豬用玉米、大豆粕飼料 (CP 12%) 之必需氨基酸含量
要求量% |
飼料中含量% |
要求量 充足率% |
|
arginine |
0.20 |
0.77 |
385 |
histidine |
0.18 |
0.30 |
166 |
isoleucine |
0.50 |
0.58 |
116 |
leucine |
0.60 |
1.31 |
218 |
lysine |
0.70 |
0.50 |
71.4* |
methionine+cystine |
0.45 |
0.48 |
106 |
phenylalnine+tyrosine |
0.70 |
1.24 |
177 |
threonine |
0.45 |
0.54 |
113 |
tryptophane |
0.12 |
0.10 |
83* |
valine |
0.50 |
0.59 |
118 |
表5-7 第1及第2限制氨基酸的添加效果
飼 料 |
增重量 kg/日 |
1.玉米 + 大豆粕飼料 (CP 12%) |
0.40 |
2.飼料1 + 離氨酸 |
0.50 |
3.飼料1 + 色氨酸 |
0.30 |
4.飼料1 + 離氨酸 + 色氨酸 |
0.60 |
在表5-6之飼料內添加第1限制氨基酸的離氨酸時成績獲得改善,但只添加第2限制氨基酸的色氨酸時成績惡化。同時添加第1及第2限制氨基酸時成績最佳。
此種意義之不平衡的場合,即添加第2限制氨基酸的場合,血液中之第1限制氨基酸的濃度減少,且添加之第2限制氨基酸的濃度顯著增大。此種血中游離氨基酸濃度會使飼料攝取量低下。血中游離氨基酸濃度的變動能被腦檢出,由腦而來的信號引起飼料攝取量的低下。此種信號強力而優先度高,在自主的飼料選擇的實驗方面,將不平衡飼料與無蛋白質飼料並列的場合,動物選擇無蛋白質飼料。
而且,此種飼料攝取量的低下似不會帶來其他生理的障礙,行飼料之強制給與時能正常的成長,不會帶來任何障礙。不平衡的場合,結果使第1限制氨基酸的血中濃度減少,使成長低下,故結果與單一氨基酸缺乏的場合很相似。 而且,胰島素的注射,寒冷下之暴露等,會減少或解消飼料之氨基酸不平衡的影響。
4. 氨基酸之拮抗作用
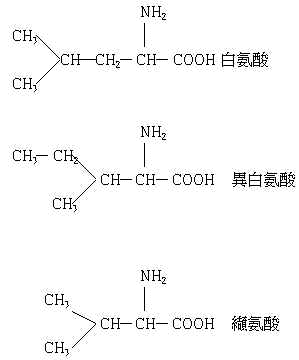
不平衡是任何必需氨基酸為第1或第2限制氨基酸時亦會引起的現象,而氨基酸的拮抗作用則是在化學構造相關連的氨基酸之間引起的現象。此種拮抗作用主見於離氨酸與精氨酸、白氨酸與異白氨酸、纈氨酸之間。拮抗作用的場合,是這些氨基酸一方增量時,另一方的要求量增加。
5. 分岐鍵氨基酸的拮抗作用
白氨酸的過剩特別使異白氨酸及纈氨酸的要求量顯著增加。分岐鍵氨基酸之拮抗作用的機構尚有很多地方不明瞭,但飼料攝取量的低下則大部分與此有關聯。
而且,給與過剩之白氨酸時血中之異白氨酸及纈氨酸的濃度顯著降低。白氨酸之過剩不太大時經過一定的期間後即能適應,飼料攝取量及成長均恢復正常。Smith等 (1978) 就雛雞、老鼠等發現白氨酸過剩時會促進異白氨酸在體內的分解。
6. 離氨酸.精氨酸的拮抗作用
鹼性氨基酸之細胞膜的通過部位是共通的。在離氨酸.精氨酸拮抗作用上,離氨酸的過剩似不會妨礙消化管之吸收精氨酸,但血中之離氨酸濃度高時腎臟之尿細管內精氨酸的再吸收減少。雛雞方面飼料中的離氨酸過剩時尿中之精氨酸排泄量增加。
精氨酸之代謝有兩點受到離氨酸過剩的影響,其一是在與從精氨酸形成肌酸 (creatin) 的第一階段有關連的酵素活性因離氨酸過剩而低下。由此而使肌酸的形成低下。給與離氨酸過剩之飼料的雛雞若在飼料中添加肌酸時能促進成長。
其二是在從精氨酸形成鳥氨酸 (ornithine) 之際 arginase活性在離氨酸過剩的場合提高,離氨酸的過剩使腎臟的arginase活性提高數倍,被分解為鳥氨酸及尿素的精氨酸量顯著增多。據Austic的報告,離氨酸過剩的場合,在此種經路下被分解的精氨酸達到攝取量的30~40%。
似此,肌酸形的不足及精氨酸分解的促進除了引起精氨酸的不足外,飼料攝取量的低下亦會引起成長的低下。
7. 精氨酸、甘氨酸與甲硫氨酸、甲硫氨酸與羥丁氨酸的相互作用
Fuller等 (1971) 在雛雞身上發現出精氨酸,甘氨酸及甲硫氨酸的相互作用。在甲硫氨酸稍微不足的玉米.大豆粕飼料內添加過剩之精氨酸時成長顯著低下。加入甘氨酸時其低下更為增大,加入甲硫氨酸或其他甲基供與體時能預防成長的低下。
過剩之精氨酸的添加使肌酸的排泄及組織的肌酸含量增大,反之,肌酸的添加能預防精氨酸引起成長的抑制。精氨酸之過剩引起甲硫氨酸要求量的增大,係為了因應增大之肌酸合成所需要的甲基。
關於甲硫氨酸與羥丁氨酸的相互作用Girard-Globa等曾有報告。過剩之甲硫氨酸的添加二次性的引起羥丁氨酸的缺乏,或引起成長的低下。
過剩之甲硫氨酸添加,使血漿及組織的羥丁氨酸濃度顯著低下,此點在老鼠方面已有發現。
飼料營養雜誌第二冊合訂本
1987年元月號至1987年十二月號
飼料營養:1987年第六期(79)
Copyright © 1998 ![]() 茂群峪畜牧網.
本網站圖文係屬茂群峪有限公司,內文之版權為該雜誌社所
茂群峪畜牧網.
本網站圖文係屬茂群峪有限公司,內文之版權為該雜誌社所
有,非經本公司及該雜誌社正式書面同意,不得將全部或部分內容,
轉載於任何形式媒體
※ 最佳解析度 800x600
Copyright © 1998
MiobufferCo., Ltd. All rights reserved.
Unauthorized copying and reproduction is prohibited. All trademarks property of their
respective holders.