臺灣土雞血液型之探討(上)
摘要
為明瞭現存臺灣土雞之品種特徵,作為進一步利用該珍貴種原基因的依據;乃利用六個不同品種(臺灣土雞、單冠白來亨雞、烏骨雞、無鱗雞、名古屋種及橫斑蘆花)計十二個品系,總數419隻雞的血球與血清(或血漿樣本),以凝集反應測試法與聚丙烯醯胺膠體電泳法分別進行紅血球表面抗原型B系統與血清蛋白型(包括血清前白蛋白、白蛋白、後白蛋白與鐵白蛋白)分析,期瞭解各品系之遺傳變異。結果顯示除後白蛋白在各品種、品系雞隻無多態性變異外,餘四個基因座均呈現多態性變異;且紅血球B系統基因座之異質結合率較其餘四個血清型基因座的平均異質結合率為高(0.603 vs 0.402)。此外,臺灣土雞J品系之前白蛋白─1(prealbumin-1)與鐵合蛋白(transferrin)出現異於其他品種或品系之表型(BB,BC與BB,BC);而單冠白來亨雞則在白蛋白(albumin)出現特殊表型AB。同時,由遺傳類緣關係與遺傳距離之分析結果發現,除土雞J及11品系與其它品系雞隻遺傳距離較遠外,餘十個品系雞隻可劃分為A與B二個集團。其中A集團包括土雞品系7、12、K、烏骨雞與無鱗雞;而B集團則由單冠白來亨雞的二個品系E、P,名古屋、土雞品系9與橫斑蘆花雞構成。(關鍵語:台灣土雞、血型、多態型)
緒言
幾世紀以來,畜牧生產在追求高度商業利益的原則下,僅具高生長與繁殖效率或其他重要經濟性能之畜禽品種得以留存,因而導致許多家禽品種已瀕臨絕跡,造成全世界家畜禽遺傳變異性的大量減少。為了突破目前經濟性能改良的瓶頸及對〞抗病育種〞的強烈訴求,對本地品種基因的尋回、保存與利用,已成為世界各國畜產業的重大課題(戴,1990),而畜產種原的保存與利用方式的選擇則關係著臺灣畜產文化之演變。臺灣省畜產試驗所及附屬單位與大學畜牧系近年來開始著手的各項本地種原保存計畫乃以下列四種方式進行:(1)小族群繁殖保種,(2)冷凍保存其生殖細胞及胚,(3)與外來品種進行雜交選育,(4)收集其生長、繁殖及生活資料,截至目前已有相當的成果(吳,1990)。
臺灣地區擁有不少本地種家畜禽種類,諸如黃牛、水牛、山羊、桃園種豬、小耳種豬、菜鴨、中國鵝以及土雞等;這些品種對臺灣地區環境的高度適應性所表現出來的特微,如耐粗性、抗熱性、多產性與抗病性等均為改良進口種畜禽的重要基因來源(戴,1990)。土雞為臺灣本地種雞,也是最重要的肉用雞之一(李與黃,1988),其可口性業已經消費大眾所肯定;鉛色腳脛、有色羽毛、活躍性強、生產淺褐色蛋與性強為其共同點。同時,較佳的抗熱與抗病力(包括白冠病、馬立克病、新城雞瘟與球蟲病;Lee and Huang,1989),則是該品種十分重要的優點。至於本地雞之來源已不可考(李,1986)。根據淺野(1944)所載,本地雞(臺灣在來雞)之品種甚多,毛色複雜,無固定品種之特徵,體質強健,就巢性強,適應環境之能力較任何雞隻為強,且耐粗。成熟體重:公雞約為2.0公斤;母雞約為1.5公斤。年產60~80個蛋,平均蛋重約37~45公克,不適蛋用,宜於肉用。臺灣民國飼養者,95%以上屬於這類雞種。自明代以來,先民帶來大陸雞,日據時代又曾引進來亨雞(Leghorn)、名古屋種(Nagoya)、三河種、洛島紅(Rhode Island Red)、橫斑蘆花雞(Barred Plymouth Rock)、澳洲黑(Australorp)、鬥雞與矮雞等雞種。光復後,政府亦曾自美國引進紐漢西種(New Hampshire)。因此,所謂〞真正土雞〞,意指「本地雞」,但可能混有本地與外來雞種血統,而經農家飼養適應本地生長環境且受農民喜好的雞種(李,1986);故其品系之混雜當可想而知。因此,和其他畜禽一樣,確立其品種特徵作為純種與雜交個體鑑別的依據,乃當今要務。應用遺傳變異(genetic variation)分析比較族群間之類緣關係為族群遺傳學研究之重要工具之一,而諸多分析遺傳變異方法中,由於血型(如紅血球表面抗原型與血液 化多態型),蘊藏豐富遺傳變異訊息,故普遍應用於族群構造的分析(Hcdrick, 1985)。有關雞血型方面的研究很多,其大多著重於遺傳標記、與經濟形質的關連性(Rendel, 1968;Rashid, 1982)、與組織適合性和抗病性之相關(Hasen et al., 1967),或血型族群遺傳特性之探討(橋口等,1981)。本篇旨在根據品種或品系間血型基因座上對偶基因頻率之差異,分析比較各品種、品系雞隻間遺傳組成之變異性(variability)和類緣關係(similarity),期為臺灣土雞未來遺傳育種研究上之參考依據。
材料與方法
一、試驗動物
本試驗供試之419個血樣(血球、血清或血漿)係分別採自臺灣省畜產試驗所及國立中興大學畜產學系雞群,其各品系來源及採樣數目如表1所示。
表1. 供試雞隻品種、隻數與來源
| 品種/品系 | 隻數 | 保種單位 | 來源 | 備註 | 文獻 |
| 臺灣土雞 7 9 11 12 J K |
41 54 28 43 45 17 |
臺灣省畜產試驗所 〞 〞 〞 國立中興大學 〞 |
臺南縣 臺東縣 嘉義縣 臺中縣 嘉義縣竹崎鄉 新竹縣關西鄉 |
民國74年收集 〞 〞 〞 民國73年收集 〞 |
鍾等, 1990 〞 〞 李與黃, 1988 |
| 單冠白色來亨雞 E P |
40 42 |
臺灣省畜產試驗所 〞 |
本省各地 〞 |
65年收集,源自同一族群 著重產重選拔 著重產蛋數選拔 |
姜等,1984
|
| 烏骨雞 | 30 | 臺灣省畜產試驗所 | 74年自本省民間雞場購入 | ||
| 無鱗雞 | 21 | 臺灣省畜產試驗所 | 美國加州大學 | 78年引種,由紐漢西突變而來 | |
| 名古屋 | 41 | 國立中興大學 | 日本 | 民國76年引進 | |
| 橫斑蘆花 | 17 | 臺灣省畜產試驗所 | 美國 | 民國49年引進 |
二、血樣收集
將絕食12小時以上之雞隻固定,以NO.23(1")針頭自雞隻翅部內側之肱靜脈管(brachial vein)採取約5~10ml的血液,經離心分離為血球與血清(或血漿)後,進行紅血球表面抗原B系統與血清蛋白型分析。
三、試驗方法:
(一)紅血球表面抗原型B系統(Ea-B)之分析:以臺灣省畜產試驗所開發且經與國際標準抗血清同定(identify)的單株抗體(monoclonal ant ibody)B2、B24、Bu、B19與B14計五株(劉,1991)與各供試雞隻之紅血球以96孔(12×8)的微量滴定盤(Micro platte, Greiner, U-form)進行凝集反應測試。
(二)血清蛋白型分析:分析項目包括前白蛋白─1、白蛋白、後白蛋白─A及鐵合蛋白等四種。由於各蛋白型之解析度將受蛋白質濃度之影響,故冷凍血清經解凍後,輔以追蹤染劑作適當之稀釋;並以修正Laemmli(1970)非連續性緩衝系統聚丙烯醯胺膠體電永法(discontinuous buffer system polyacrylamide gel electrophoresis ;以下簡稱PAGE)進行分析。
(三)統計分析
首先考慮品系之第 i個體第 j個基因座具n個對偶基因(K=1,2,……,n),則可組成C(n+1,2)種可區分表型,其中n種為同質結合體,C(n,2)種異質結合體;則依哈第─溫柏定律(Stern,
1943),其第K個對偶基因頻率與變方之估計值(![]() )為
)為
上式中,NKK為帶第K個對偶基因之同質結合體個數;NKK為帶第K個對偶基因之同質結合體個數;N則為總個體數。
應用前述基因頻率估計值,吾人可得第
i個體第 j個基因座之哈第─溫柏異質結合率(Hardy-Weinberg Heterozygosity)估計值,Hij,如下: 並依Nei and
Roychoudhury(1974)公式,可進一步求得抽樣變方(sampling variance)估計值,V(Hij),
並依Nei and
Roychoudhury(1974)公式,可進一步求得抽樣變方(sampling variance)估計值,V(Hij),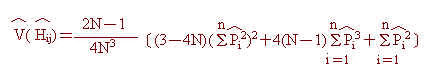
但若樣本族群數目小於30時,以上二式均需經過(2N/2N-1)因子予以矯正(Nei and Roychoundhury, 1974)。
因此,本研究中各品系之平均異質結合率(H)與其抽樣變方(V(H))可估算如下:
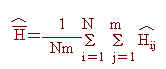
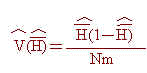
其中除m為所偵測之基因座總數外,餘各項定義均如前述。同時,為瞭解各基因座之各對偶基因頻率之平均性,乃利用同質結合率之倒數,ne,評估之,意即ne之估計值,︿ne,可估算如下:
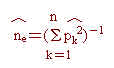
就某一個基因座而言,上式同時考量總對偶基因數目(n)與各對偶基因頻率估計量(︿pk),其通稱為有效基因數目(the number of effective alleles, Marshall and Brown, 1975)估計值。
多型基因座比例(proportion of polymorphic loci)之估算為本研究中用以評估各品系遺傳變異之另一方法,其乃是計算多型基因座數目佔所有被偵測基因座數目之百分比,估計式與相對應變方估計值如下:
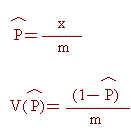
式中,x為具有多型性基因座之數目;m則為所偵測基因座數目。由於本研究中,各品系之測試個體數目均小於100,故多型基因座係指最常出現對偶基因頻率小於0.95者。
本研究中各品系間遺傳類緣關係之分析,乃是應用估算所得五個基因座上各對偶基因頻率估值,並採用Nei(1972)所發展遺傳一致值(genetic identity;I11')與遺傳距離(genetic distance;D11')估算公式而得。首先令︿Pk1與︿Pk1分別為品系1與1'之第 j個基因座( j=1,2……,5)之第K個對偶基因頻率估計值,且
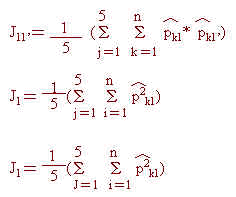
則可得I11'與D11'之估計值如下:
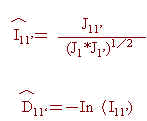
俟完成各品系間遺傳距離之估算後,再配合群體分析(cluster analysis)之應用(Sokal and Sneath, 1963),將遺傳距離最小二個品系聚集成一單位,並以算術平均數法求其與其他品系間之距離;週而復始逐步建立所有品系間之遺傳距離。
結果與討論
一、紅血球表面抗原型B系統(Ea-B)
以畜試所製備而得的B2、B24、Bu、B19與B14等五株單株抗體(monoclonal antibody)測定六個品種(12個品系)供試雞隻之376個血樣之紅血球表面抗原型B系統,發現各品系(strain)雞隻在這五種紅血球表面抗原型之基因頻率分布不盡相同(表2)。然因單株抗體僅有五株,故無法確定同質結合體;並將未測出基因型歸於其他(others)。因而有低估該五種對偶基因頻率的傾向,意即所得到之基因頻率應為保守估值。同時,若以種原收集之地域觀之(表1與表2),品系11與J雖為畜試所與中興大學分別於民國74年與73年同自嘉義地區收購而得,然其紅血球表面抗原B系統基因座之最主要對偶基因並不相同。究其原因,J系為自嘉義竹 崎1公,6母繁殖而得;而品系12則為畜試所自嘉義縣地區購得之46隻土雞,經全同胞近親配種而選留大家族(large family size),故其基因頻率不同,雖可能係因採樣差異,亦可能為選拔所造成者。
表2. 不同雞種之血型B系統基因頻率
| 品種/品系 | 隻數 | B基因座基因頻率 | |||||
| B2 | B24 | Bu | B19 | B14 | others | ||
臺灣土雞 |
|||||||
| 7 | 36 | .056±.027 | .125±.039 | 0 | .306±.054 | .125±.039 | .388 |
| 9 | 50 | .150±.036 | 0 | .090±.029 | .020±.014 | 0 | .740 |
| 11 | 25 | .060±.034 | .040±.028 | .320±.066 | .040±.028 | .020±.020 | .520 |
| 12 | 40 | .125±.037 | .050±.024 | .075±.029 | .238±.048 | .238±.048 | .275 |
| J | 45 | .400±.052 | .333±.050 | .056±.024 | 0 | .100±.032 | .111 |
| K | 12 | .167±.067 | .208±.083 | .042±.041 | 0 | .167±.076 | .417 |
單冠白色來亨雞 |
|||||||
| E | 40 | .113±.035 | 0 | 0 | .300±.051 | 0 | .588 |
| P | 39 | 0 | 0 | 0 | .295±.052 | 0 | .705 |
| 烏骨雞 | 21 | 0 | 0 | .071±.004 | .333±.073 | 0 | .595 |
| 無鱗雞 | 21 | .214±.063 | 0 | .143±.054 | .405±.076 | 0 | .238 |
| 名古屋 | 41 | .183±.043 | 0 | .012±.012 | .207±.045 | 0 | .598 |
| 橫斑蘆花雞 | 6 | 0 | 0 | .250±.125 | 0 | 0 | .750 |
此外,在單冠白來亨雞方面,E與P系出現B2基因頻率分別為0.113與0,此可能因該二品系之長期選拔目標不同,而造成該二個品系之紅血球表面抗原型B系統基因頻率分布呈顯著性差異(P<0.05)。再者,前述結果與Abplanalp et al.(1985)之研究發現選拔高產蛋數會導致B2基因消失之結果一致。以土雞各品系與其他品種雞隻比較,發現僅土雞具有B24與B14對偶基因,而中興大學所保種之土雞J與K品系和畜試所四個土雞品系,則因前者未檢出B19型而有所不同。
二、血清蛋白型
(一)血清前白蛋白─1型(Pa-1):以PAGE法檢測六個雞種之血清前白蛋白─1型,發現有三個對偶基因(A,B,C)控制其多態型表現。六不同品種雞之Pa-1外表型中,除名古屋種全部為AC型外,餘均有二種或以上的外表型檢出,且多屬於AB與AC二型(表3)。又BB型僅出現於品系J;而BC型在品系12只出現一例,然在品系J中發現有20例之多。綜合言之,各品系之雜合子型個體(heterozygote)均較純合子個體(homozygoye)為多;又純合子之AA與CC型在本各試驗中均未檢出,其可能原因有三:(1)採樣數不足,(2)採樣誤差,(sampling error)或(3)純合子個體胚胎早期死亡所致;然以x2 test(P<0.01)得知,應非採樣數小或採樣誤差所造成者;至於真正原因,仍需進一步之探討。各品系之Pa-1型基因頻率如表3所示。在單冠白色來亨雞方面,不論著重蛋重抑或產蛋數之選拔品系,其Pa-1之外表型及基因頻率的分佈均無顯著差異(P>0.1);此現象於白蛋白與鐵合蛋白二基因座亦如是,顯示單冠白來亨雞在歷經15年之產蛋數與蛋重之選拔後,改變了紅血球表面抗原B系統基因座的基因頻率;但對諸如血清前白蛋白─1、白蛋白與鐵合蛋白等血清蛋白型之表型與基因頻率,則未發生影響。
表3. 不同雞種之血清前白蛋白─1表型與基因頻率
| 品種/品系 | 隻數 | 表型 | 基因頻率 | |||||
| BB | AB | AC | BC | Pa-1A | Pa-1B | Pa-1C | ||
臺灣土雞 |
||||||||
| 7 | 38 | 0 | 3 | 35 | 0 | .500±.057 | .040±.022 | .461±.057 |
| 9 | 54 | 0 | 22 | 32 | 0 | .500±.048 | .204±.039 | .296±.044 |
| 11 | 25 | 0 | 11 | 14 | 0 | .500±.071 | .220±.059 | .028±.063 |
| 12 | 41 | 0 | 15 | 25 | 1 | .488±.055 | .195±.044 | .317±.050 |
| J | 38 | 9 | 5 | 4 | 20 | .118±.037 | .566±.057 | .316±.053 |
| K | 17 | 0 | 3 | 14 | 0 | .500±.086 | .088±.049 | .412±.084 |
單冠白色來亨雞 |
||||||||
| E | 35 | 0 | 3 | 32 | 0 | .500±.060 | .043±.024 | .457±.060 |
| P | 42 | 0 | 2 | 40 | 0 | .500±.055 | .024±.017 | .476±.054 |
| 烏骨雞 | 30 | 0 | 3 | 27 | 0 | .500±.065 | .050±.028 | .450±.064 |
| 無鱗雞 | 21 | 0 | 10 | 11 | 0 | .500±.077 | .214±.063 | .286±.070 |
| 名古屋 | 31 | 0 | 0 | 31 | 0 | .500±.063 | 0 | .500±.063 |
| 橫斑蘆花雞 | 17 | 0 | 1 | 16 | 0 | .500±.085 | .029±.029 | .470±.086 |
(二)血清白蛋白型(Alb):血清白蛋白的次級帶(minor band)的多態型亦由三個對偶基因(A,B,C)控制其表現(表4)。在表型方面,純合子AA型在供試各品系雞隻均未檢出;BB型除於土雞9、11與12品系偶有發現,且在無鱗雞檢測到一例之外,其他各品系殆無BB型之檢出;而CC型之個體除在橫斑蘆花雞未檢出外,餘各品系均有檢出之例。雜合子型方面,除AB型僅出現在單冠白色來亨雞外,其他雜合子型AC與BC型,則在各品系中均佔有較多的個體數目 。綜觀之,土雞、烏骨雞、無鱗雞以及名古屋雞之血清白蛋白次級帶表型之多態型表現,以BC型居多;而單冠白色來亨雞與橫斑蘆花雞則以AC型為主。整體而言,雜合子之總個體數遠較純合子為多,與血清前白蛋白─1型結果相同。此外,AlbA、AlbB與AlbC基因出現頻率最高的雞隻品種分別為橫斑蘆花雞(0.438)、名古屋雞(0.371)與土雞K品系(0.667)。
表4. 不同雞種之血清白蛋白(Alb)次級帶表型與基因頻率
| 品種/品系 | 隻數 | 表型 | 基因頻率 | ||||||
| AB | BB | CC | AC | BC | AlbA | AlbB | AlbC | ||
臺灣土雞 |
|||||||||
| 7 | 41 | 0 | 0 | 8 | 9 | 24 | .110±.035 | .293±.050 | .598±.054 |
| 9 | 54 | 0 | 4 | 11 | 16 | 23 | .148±.034 | .287±.044 | .565±.048 |
| 11 | 29 | 0 | 1 | 10 | 13 | 5 | .224±.055 | .121±.043 | .655±.062 |
| 12 | 43 | 0 | 4 | 7 | 12 | 20 | .140±.037 | .326±.051 | .535±.054 |
| J | 44 | 0 | 0 | 12 | 11 | 21 | .125±.035 | .239±.045 | .636±.051 |
| K | 15 | 0 | 0 | 5 | 3 | 7 | .100±.055 | .233±.077 | .667±.086 |
單冠白色來亨雞 |
|||||||||
| E | 35 | 1 | 0 | 1 | 20 | 13 | .300±.055 | .200±.048 | .500±.060 |
| P | 41 | 5 | 0 | 1 | 20 | 15 | .305±.051 | .244±.047 | .451±.055 |
| 烏骨雞 | 30 | 0 | 0 | 6 | 8 | 16 | .133±.044 | .267±.057 | .600±.063 |
| 無鱗雞 | 21 | 0 | 1 | 5 | 5 | 10 | .119±.050 | .286±.070 | .595±.076 |
| 名古屋 | 31 | 0 | 0 | 2 | 6 | 23 | .097±.038 | .371±.061 | .532±.063 |
| 橫斑蘆花雞 | 16 | 0 | 0 | 0 | 14 | 2 | .438±.088 | .063±.043 | .500±.088 |
(三)血清後白蛋白─A型(Pas-A):後白蛋白於雞隻血液型中,依其異動度(mobility)的大小可分為二個區域,分別由不同的基因座控制其表現,且基因座上的對偶基因均為簡單的顯隱性關係。依Kuryl and Gasparska(1976)研究報告,Pas-A出現於所有公雞與大部分的母雞,而Pas-B則僅出現於產蛋期母雞;故本試驗所偵測者為易動度較大的後白蛋白─A型。結果發現所有供試雞隻,無論品系與性別,均未出現任何隱性純合子表型,意即均可檢視到Pas-A帶。此與Tanabe and Mizutani(1980)及Okada et al.(1984)報導之日本烏骨雞與名古屋雞種,呈多態型變異的結果不同,或因PAGE具有較佳之解析力有以致之。同時,該基因也是本試驗中所探討五個基因座中唯一於各品系均不呈多態性變異者(表5)。
表5. 不同雞品種之五種蛋白型基因座多態性a
| 品種/品系 | 蛋白型基因座 | ||||
| Ea-B | Pa-1 | Alb | Pas-A | Tf | |
| 臺灣土雞 | |||||
| 7 | P | P | P | ─ | P |
| 9 | P | P | P | ─ | P |
| 11 | P | P | P | ─ | P |
| 12 | P | P | P | ─ | P |
| J | P | P | P | ─ | P |
| K | P | P | P | ─ | P |
| 單冠白色來亨雞 | |||||
| E | P | P | P | ─ | P |
| P | P | P | P | ─ | P |
| 烏骨雞 | P | P | P | ─ | ─ |
| 無鱗雞 | P | P | P | ─ | P |
| 名古屋 | P | P | P | ─ | P |
| 橫斑蘆花雞 | P | P | P | ─ | P |
a因個體數目小於100,故最常出現之對偶基因頻率<0.95時,則為具多態型基因座。
飼料營養雜誌(p.11~20)─薛佑玲.九三年第五期