離胺酸與豬禽營養
1. ADM一一面向世界的超級市場
ADM是世界上最大的農產品加工和處理公司。該公司在世界各地擁有135家生產廠家,每年向世界源源不斷地提供越來越多的糖類、植物油、麵粉、澱粉、燃料用乙醇、動物飼料等等。
該公司同時也生產附加價值較高的產品,用於食品、飼料和其它各行工業。這些產品源於植物蛋白、油或玉米中提取的糖類。
ADM公司使用世界上最先進的發酵技術生產離胺酸、蘇胺酸和色胺酸。當然其它產品亦在生產範圍之內,如:檸檬酸、乳酸、葡萄糖鈉鹽和澱粉分解𠕇。該公司不斷投資於研究和開發,以擴大桿菌滛和生物素的生產規模。
不僅限於農產品的加工處理,ADM公司的經營範圍還包括穀物買賣,遠洋和陸地運輸以及全球性的市場投資。該公司是農產品及其副產品轉化成肉類、奶類、蛋類和其它食物的中轉站。
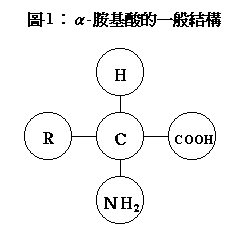
胺基酸是簡單的有機低分子物,通常被稱為蛋白質的組成基石。所有蛋白質均為胺基酸殘基相連而成。組成蛋白質的胺基酸全是α型,至少含有一個羧基和胺基,後者結合在α─碳原子上。α─胺基酸的基本結構見圖-1,蛋白質中常見的胺基酸有20種,它們的不同之處在於"R"位上的基團和側鏈。
胺基酸在自然界中普遍存在,其歷史在地球上已有幾十億年,然而直到1806年人們才發現第一個胺基酸。最後一個被發現的蘇胺酸(1935)。離胺酸是DRECHSEL於1889年從酪蛋白中首次分離出來的。
在自然界中,所有的胺基酸都能被微生物和綠色植物合成。這是它們利用氨的主要途徑。相反,人類和動物合成胺基酸的能力極為有限。大多數動物只能合成蛋白質存在的二十多種α─胺基酸中的十種,其它十種胺基酸常被稱為"必需胺基酸",這些胺基酸只能從食物中攝取。離胺酸是動物營養中最重要的必需胺基酸之一。
盡管離胺酸在生化工業界少量生產已有幾十年歷史,然而進入大規模的商業化生產則始於60年代,這個過程涉及到某些微生物的分離和培養,利用這些培養好的微生物進行大規模的發酵,將葡萄糖轉化成離胺酸。這種生物高技術在效率方面得到長足的進步,使得離胺酸的世界產量在1975-1980年提高了整整三倍。1985又比1980年提高了兩倍。
2. 離胺酸的一般特性
離胺酸的幾種特性見表-1。離胺酸的結構尤其有趣,因為它具有兩個胺基。連在第五個碳原子上的胺基鹼性極強,只有在PH值很高的條件下才失去質子。所以離胺酸通常被認為是鹼性的。連在第五位碳上的自由胺基有很強的化學反應性,因而此胺基較易與其它化合物產生化學上的結合,特別是與還原性的碳水化合物。這種結合產物的化學反應過程通常被稱為"褐變反應"或"米勒反應"。這些化學上的聯繫使離胺酸失去其生物效果。
純的或游離的離胺酸的吸濕性很強。因此,商業化生產的離胺酸通常以離胺酸鹽(鹽酸)的形式出現。這樣能使離胺酸比較穩定且幾乎具有無色無味的性質。這種形式的離胺酸的一些特性列在表-1。

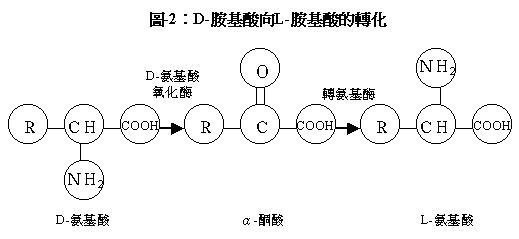
離胺酸能以L-或D-的立體異構兩種形式存在。但只有L-型存在於生物的蛋白質中。D-胺基酸能被轉化成L-型胺基酸。這個過程包括兩步:D-胺基酸經氧化變成它(們)的α-酮酸;後者經轉胺基作用產生L-型(見圖-2)。遺憾的是動物不具備第一步反應中的D-胺基酸氧化𠕇來執行它的第一步轉變。因此,D-離胺酸沒有生物活性,但是,所有商業化生產的離胺酸均為均為L-型離胺酸。
3. 離胺酸的生物學作用:
離胺酸的幾個重要的生物學作用列在表-2和圖-3。在消化過程中,結合在飼料蛋白質內的離胺酸被釋放為游離離胺酸。離胺酸的代謝途徑有多種。最重要的是作為必需胺基酸用於各種蛋白質的合成,特別是肌肉蛋白和𠕇。離胺酸是肌肉蛋白質含量最多的胺基酸之一,同時也是幾種滛類激素的重要組成成分。
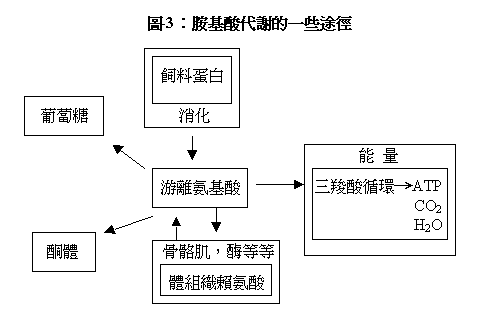
表2、離胺酸在生物體中的作用:
蛋白質合成─ ─骨骼肌、𠕇、滛類激素𠕇等 糖原合成 酮體合成 肉毒鹼的前體─脂肪酸代謝 能量來源 酸/鹼平衡 離胺酸/精胺酸拮抗 |
離胺酸是少數幾種既能產糖又能生酮的胺基酸之一(見圖-1)。所以,當體內缺乏可利用的碳水化合物時,離胺酸經代謝後能產生糖或酮體。對於攝食量極低(如斷食)和某些類型的糖尿病患者,這一途徑能提供極其重要的能量來源。離胺酸是肉毒鹼的前體,後者對於脂肪代謝極為重要。因為它是位於線粒體膜上的一種𠕇的重要組成成分,這種𠕇能使長鏈脂肪酸通過線粒體膜而被氧化產生能量。
前面已提到多餘的離胺酸能被氧化當作能量來利用,其ME值達4600千卡/公斤。離胺酸的非氮部分能進入三羧酸循環(TCA),進而氮化產生水、二氧化碳和能量(ATP)(見圖-3)。然而,以這種形式提供能量其效率很低。因為,離胺酸的含氮部分必須轉化成尿酸和尿素等而排泄,而此過程需要消耗能量。
3.1 離胺酸和精胺酸的拮抗作用:
離胺酸與精胺酸的拮抗作用是胺基酸拮抗中最為熟知的一種。這種拮抗對於家禽來說特別重要。飼料中過多的離胺酸造成腎中精胺酸分解𠕇活力升高,從而使更多的精胺酸降解。如果飼料中精胺酸的含量得不到相應的提高,過多的離胺酸會造成家禽生長速度下降。但只要飼料中精胺酸含量相應提高,這一不良作用能被減輕。因此,飼料中離胺酸的含量不應超過精胺酸含量的20%。不過,這種情況在實際飼餵條件下很難出現。相反,過多的精胺酸同樣能拮抗離胺酸,特別是當飼料中蛋白質和離胺酸含量很低時。不過相比之下,精胺酸對離胺酸的拮抗作用比離胺酸對精胺酸要小。離胺酸與精胺酸拮抗的作用對豬來說不是很重要,因為豬對精胺酸的需求量很低,而且精胺酸能在尿素循環過程中合成。
如果精胺酸含量充足的話。離胺酸的致毒作用很小,對於家禽來說,即使飼料中含量高達2.4%,也不至於產生任何不良效果。
3.2 蛋白質和離胺酸的周轉:
在體內,蛋白質的合成和降解過程是同時發生的,位於體內不同的各種胺基酸代謝池也不斷輸出胺基酸用於蛋白質的合成。同時在蛋白質的降解過程中,胺基酸則不時進入體內各種胺基酸代謝池,這一動態過程被稱為蛋白質的周轉。這裡值得重視的是大約80%的離胺酸和其它各種胺基酸通過蛋白降解釋放後又被重新用於蛋白質的合成。從蛋白降解後,只有20%的胺基酸經尿液排出。用於生長動物的飼料必須提供足夠的胺基酸來彌補這20%的損失,與此同時,還得有足夠的數量用於組織生長。
3.3 飼餵頻率對晶狀離胺酸在體內代謝和利用的影響:
早期做的研究工作顯示:當豬每日只飼餵一次時,飼料中添加的晶狀離胺酸的利用效率只相當於每日多次飼餵效率的50-67%。有人認為這種利用效率不高是因為從腸道吸收晶狀離胺酸的速度要比日糧中結合於蛋白質中的胺基酸吸收要快得多。因此,在採食後的某一時期離胺酸的相對濃度比其它用於蛋白質合成的胺基酸要高得多,這過高部分的離胺酸因其它胺基酸濃度的相對不足被降解了,而不是用於合成蛋白質。當動物每日多次飼餵就能保證所有的胺基酸濃度比較適合於蛋白質的合成。在這種情況下,離胺酸的利用效率就高。
美國伊利諾大學最近的研究結果表明,當雞每日飼餵兩次(間隔12小時),每次餵飼時間為1小時,晶狀離胺酸的利用效率與自由採食效率很相近。飼料中總離胺酸的利用效率沒有因為添加晶狀離胺酸而降低,以上結果和其它研究表明:動物可能在離胺酸攝入不足的情況下仍能保持離胺酸的濃度。當動物採食不含蘇胺酸的飼料時,其肝和肌中游離蘇胺酸濃度下降的程度要遠遠超過當動物採食不含離胺酸飼料時其肝和肌肉中游離離胺酸濃度下降的程度。此外當動物採食不含離胺酸飼料時,其體重下降的程度遠不如當動物採食缺乏其它必需胺基酸飼料時體重下降的程度。這些結果表明由於離胺酸缺乏,所產生組織蛋白的降解和自由胺基酸代謝池中各種胺基酸的降解,在程度上均不如其它必需胺基酸缺乏所產生的程度大。
伊利諾大學在雞、以及最近在豬方面所做的研究工作表明;在豬和雞每日飼餵兩次的情況下,添加的晶狀離胺酸其利用效率與多次飼餵沒有太大差別,與其它胺基酸比較,離胺酸的生物利用效率不會因每日飼餵次數而受到任何不良影響。這與前面提到的有不同之處。
4. 胺基酸需要量:
4.1 一般性原理:
豬禽的胺基酸需要量列於表-3和表-4,蛋白質需要量也列在表中作為參考,豬禽對蛋白質的需要量通常被認為,實際上是指對組成蛋白質中各種胺基酸的需要量,而並不是指需要蛋白本身。蛋白質需要量包括:(1)提供動物所需要的必需胺基酸,和(2)提供非必需胺基酸或胺基氮來合成這些非必需胺基酸。值得注意的是最近對低蛋白質添加胺基酸日糧的研究結果表明,家禽沒有蛋白質需要量這一嚴格概念,值得懷疑。在許多飼養條件下,低蛋白日糧中添加所有必需胺基酸和所需胺基氮後,所能產生的生產效果與高蛋白質的實用典型日糧的生長效果並不相同,後者的生長效果優於前者。伊利諾大學最近的研究工作還證明的蛋白質含量19%的日糧添加必胺基酸能維持小肉雞(0-3週)的最佳生長效果,而在日糧含蛋白低於16%的情況下,不管添加多少必需胺基酸,所能產生的效果遠不如前者。
表中所列出的胺基酸需要量指的是L-型胺基酸。在當飼料營養物質以百分數為計算標準的情況下,有幾種因素能間接影響這些胺基酸的需要量。這些因素與採食量有關,通過影響採食量而間接影響動物對這些胺基酸的百分需要量。胺基酸的百分需要量應根據環境的溫度、日糧能量濃度和其它任何影響採食量的因素進行適當調整。飼料中蛋白質的百分含量也能影響胺基酸需要的百分含量,這種影響同樣與採食量有關。
在幾種類型家禽中,特別是肉雞對飼料蛋白質的濃度較為敏感,蛋白質濃度增高時,其採食量相應地下降。因此,在這種情況下,胺基酸的百分濃度在蛋白質濃度高的情況下應相應提高。如果非蛋白氮含量充足且所有必需胺基酸生物效價極高,則蛋白質濃度對這些胺基酸攝入量影響不大。
表-3和表-4中所列的必需胺基酸標準應能滿足家禽和肉豬的最快生長速度(經濟效益指示沒有列入表中)。表中提供的最快生長速度所需的營養水平並不一定產生最佳經濟效益。
4.2 家禽的胺基酸需要量:
肉雞和火雞的胺基酸需要量常用飼料鳥分比和每兆卡每公斤飼料百分比的形式表達,胺基酸的濃度常根據飼料能量濃度而調整,因為後者是影響採食量的重要因素。當然影響採食量的因素還有很多。例如:肉雞採食量常隨飼料中蛋白質和胺基酸濃度的下降而增加,即使在飼料能量水平保持不變甚至增加的情況下亦如此。表-3所列的標準只是針對雄性家禽而言的。人們對雌雄分離飼養的興趣越來越高。雌性對胺基酸的需要量比雄性要低些,但目前所掌握的研究資料還無法準確地制定出與此相適應的飼養標準。四週齡以下的火雞的胺基酸需要已有很詳細的闡述。但四週齡以上的數據則很少(除離胺酸和蛋胺酸外),幾家實驗室的研究結果均證實根據表-3所配的飼料應該產生最佳生長速度。
產蛋雞的胺基酸需要量用飼料中含胺基酸的百分比計算,同時也以日採食量表示,絕大多數商業化養禽場以計算日採食量為基準,因為準確地掌握採食量對於取得最佳生長效果和最佳飼料轉換率極其重要,許多養禽場隨著產蛋雞年齡的增加和產蛋率下降而逐漸降低飼養水平。特別是後者,其目的主要是減緩蛋的體積增加率。研究結果還表明在整個產蛋週期中,最佳產蛋量(整個產蛋的重量或數量)時的胺基酸需要基本是穩定的。尚未成熟的產蛋雛雞的胺基酸需要量還未明確制定,表-3所列的標準能充分滿足在常溫下的飼餵水平。
這裡值得注意的是控制生長母雞的蛋白質和胺基酸的飼餵水平。現代品種食慾好生長快且成熟早。稍過高的飼餵水準將可能導致能量攝入過高,胴體脂肪沉積過快,而引起在產蛋周期內脫肛比例上升。
表-3 家禽的胺基酸需要
| 類別 | 粗蛋白 | 蛋氨酸 | 蛋氨酸+胱氨酸 | 賴氨酸 | 蘇氨酸 | 精氨酸 | 異亮氨酸 | 纈氨酸 | 色氨酸 |
| 肉雞: 0-21日齡1 0-21日齡2 22-12日齡1 22-42日齡2 43-56日齡1 43-56日齡2 種雞(%飼糧)1 種雞(克/日)1 |
23 6.93 20 6.25 18 5.57 14.5 22 |
.50 .155 .38 .145 .32 .114 .35 .520 |
.93 .273 .72 .255 .60 .209 .55 .820 |
1.20 .368 1.00 .318 .85 .241 .51 .765 |
.80 .241 .74 .232 .68 .191 .48 .720 |
1.44 .400 1.20 .368 1.00 .300 .74 1.11 |
.80 .245 .70 .236 .60 .205 .57 .850 |
.82 .309 .72 .282 .62 .232 .63 .950 |
.23 .073 .18 .054 .17 .050 .13 .190 |
| 火雞: 雄性:0-4周1 雌性:0-4周1 雄性:4-8周1 雌性:4-8周1 雄性:8-12周1 雌性:8-11周1 雄性:12-16周1 雌性:11-14周1 雄性:16-20周1 雌性:14-17周1 雄性:20-24周1 雌性:17-20周2 候選種雞1 產蛋雞種雞1 |
28
26
22
19
16.5 5.11 14 4.20 12 14 |
.53
.45
.38
.33
.28 .086 .23 .068 .20 .20 |
1.05
.90
.75
.65
.55 .173 .45 .136 .40 .40 |
1.6
1.5
1.3
1.0
.80 .250 .65 .195 .50 .60 |
1.0
.93
.79
.68
.59 .186 .50 .150 .40 .45 |
1.6
1.5
1.25
1.1
.95 .295 .80 .241 .60 .60 |
1.1
1.0
.85
.75
.65 .205 .55 .168 .45 .50 |
1.2
1.1
.94
.80
.70 .218 .60 .182 .50 .58 |
.26
.24
.20
.18
.15 .045 .13 .041 .10 .13 |
| 產蛋雞(來航): 產蛋雞(%飼糧)1 產蛋雞(克/日)1 |
14.5 16 |
.32 .350 |
.55 .600 |
.64 .700 |
.45 .500 |
.68 .750 |
.50 .550 |
.55 .600 |
.14 .150 |
| 未成熟產蛋型青年母雞(來航) 0-6周 6-14周 14-20周 |
18 15 12 |
.30 .25 .20 |
.60 .50 .40 |
.85 .60 .45 |
.68 .57 .37 |
1.00 .83 .67 |
.60 .50 .40 |
.62 .52 .41 |
.17 .14 .11 |
註:1. 以上所有需要量均以飼糧百分率計算並且以NRC(1984)標準為基礎。 2. 供給量以飼糧每公斤兆卡百分率計算(Waldroup. 1989)後者的數據是根據NRC(1984)和馬里蘭大學出版的數值制定的,生長肉雞的數據是以雄性肉雞測定的。 |
|||||||||
4.3 豬的胺基酸需要
表-4的標準是基於飼料中含胺基酸的百分比和日採食量,這些標準來源於兩方面:全美研究會NRC(1989)和EASTER(1989)的文獻資料。這兩組資料在胺基酸需要方面除異白胺酸和纈胺酸外基本一致。生長豬的標準可能因豬種和生理狀況而異。但如:青年母豬和公豬對胺基酸的需求比閹公豬更高,因為前者消耗的飼料較少且瘦肉率比後者高。NRC(1989)生長豬的標準是估測的,其假設條件是:生長豬所需的理想胺基酸及其間的比率在整個生長期間不變。這種理想模式主要是基於豬的維持需要和機體組織及母乳的胺基酸組成。各種胺基酸的需求量是根據它們對色胺酸的需求量比率計算出來的。對色胺酸比率最高者為離胺酸。這說明高濃度的離胺酸對瘦肉組織的生長是必需的。精胺酸的水平或比率(相對色胺酸)隨年齡的增長而逐減,因為成年豬能合成足夠的精胺酸,但小豬則不能。
妊娠母豬對胺基酸的需求是根據妊娠後期氮沉積所達到的滿意程度來制定的,哺乳豬的需求量包括維持成年母豬和支持生產足夠母乳的需求量。在成年公豬胺基酸需求量方面所做的研究工作很少,但一般認為懷孕青年母豬胺基酸標準足夠滿足成年公豬的需求量。
表-4 豬對胺基酸的需要
| 粗蛋白 | 賴氨酸 | 色氨酸 | 蘇氨酸 | 蛋氨酸+胱氨酸 | 異亮氨酸 | 纈氨酸 | |
| 生長豬: 1-5公斤(%) 1-5公斤(克/日) 4.5-9.1公斤(%) 5-10公斤(%) 5-10公斤(克/日) 9.1-20.5公斤(%) 10-20公斤(%) 10-20公斤(克/日) 20.4-54.5公斤(%) 20-50公斤(%) 20-50公斤(克/日) 54.5-屠宰體重(%) 50-110公斤(%) 50-110公斤(克/日) |
24 60 20 20 92 18 18 171 16 15 285 14 13 404 |
1.40 3.5 1.15 1.15 5.3 .95 .95 9.0 .76 .75 14.3 .60 .60 18.7 |
.20 .50 .16 .17 .80 .14 .14 1.30 .12 .12 2.30 .11 .10 3.1 |
.80 2.0 .68 .68 3.1 .56 .56 5.3 .48 .48 9.1 .40 .40 12.4 |
.68 1.7 .60 .58 2.7 .48 .48 4.6 .41 .41 7.8 .34 .34 10.6 |
.76 1.9 .70 .65 3.0 .53 .53 5.0 .52 .46 8.7 .40 .38 11.8 |
.80 2.0 .68 .68 3.1 .56 .56 5.3 .46 .48 9.1 .36 .40 12.4 |
| 種豬: 懷孕期(%)2 懷孕期(%)1 懷孕期(克/日)1 泌乳期(%)2 泌乳期(%)1 泌乳期(克/日)1 |
12 12 228 14 13 689 |
.43 .43 8.2 .60 .60 31.8 |
.08 .09 1.7 .13 .12 6.4 |
.30 .30 5.7 .51 .43 22.8 |
.23 .23 4.4 .36 .36 19.1 |
.37 .30 5.7 .39 .39 20.7 |
.40 .32 6.1 .68 .60 31.8 |
| 註:1. NRC(1988)標準 2. 供給量根據EASTER(1989)制定。 |
|||||||
4.4 胺基酸營養需要與胴體組成
最近十年,家禽和肉豬胴體中脂肪含量成了人們越來越關注的課題。少量脂肪沉積能改善肥育體況並能保證適口性,然而過量的脂肪沉積則造成繁殖功能下降。盡量減少胴體中不必要的脂肪含量現已成為重要的研究課題之一,因為從健康的角度來看,人們的消費觀念已轉向脂肪較少的肉類,而且胴體中過多的脂肪給肉類加工增加過多的不必要的處理程度。
體內脂肪的含量顯然受飼料左右,高能低蛋白飼料增加體內脂肪沉積,而低能高蛋白的飼料則增加體內瘦肉的比率。這一現象的原因主要與能量攝入有關,當飼料中蛋白或胺基酸含量低時,動物通過增加整個飼料採食量來滿足對蛋白質和胺基酸的需要,因而能量的攝入量相對提高,在這種情況下,動物的生長速度受到的影響不太,但飼料利用率則下降,體內脂肪沉積增加。有充足證據表明:為達到最佳胴體組成(低脂肪高瘦肉),機體對離胺酸的需求超過最快生長率對離胺酸的需求,在不久的將來,若豬生長激素能用於商業化動物生產,經這種激素處理後的豬其蛋白質沉積將更高,生長速度將更快,從而對離胺酸的需求量將更高。
4.5 在以玉米和大豆(餅、粕)為主要原料的日糧中,限制性胺基酸的排列順序:
由於晶狀胺基酸的大規模商業化生產,了解這些胺基酸在各種動物飼料配方中的限制性順序顯得越來越重要,有可能只用添加幾種限制性胺基酸就能取代可觀數量的飼料蛋白質原料。測定限制性胺基酸順序的方法可以是增加(加入法)或減少(遞減法)這些胺基酸在飼料中的濃度,前者主要是在低蛋白日糧中逐個加入各種胺基酸或加入幾種胺基酸。然後將這些加入的胺基酸逐個依次減少。這種方法比前者更容易且效率更佳,因為加入法通常需要很多處理組。但是遞減法有個很大的不足之處,即每減少一種胺基酸造成其它限制性胺基酸大量過剩,這部分過剩的胺基酸有可能抑制生長並且使得某種胺基酸比實際條件下更加缺乏。在這種條件下,離胺酸和精胺酸受到的影響尤其突出,原因是兩者之間的拮抗作用。因此,加入法能更加準確地測定胺基酸限制順序。
在以玉米和大豆(餅、粕)為主要原料的日糧中,各種胺基酸可能的限制順序列在表-5。很顯然,蛋胺酸在家禽日糧中是第一限制性的,離胺酸在肉雞和火雞日糧中是第二限制性的,精胺酸是第三限制性的,但精胺酸在生長肉雞日糧中(蛋白質含量16%)其限制性程度與離胺酸幾乎一樣。蘇胺酸和纈胺酸在肉雞日糧蛋白質濃度16%的情況下,其限制程度相同,屬第四限制性胺基酸。幼火雞胺基酸限制性順序目前還只停留在蛋胺酸的水平。精胺酸對肉雞的限制性程度(第三限制)遠遠超過對火雞的限制(排在最少第四以下),究其原因主要是所需蛋白質比火雞少很多,而且肉雞日糧中玉米(缺精胺酸嚴重)所佔的比例相對很大,產蛋雞的胺基酸排列順序也只停留在蛋胺酸的基礎上,當飼料含蛋白質濃度在13%的情況下,蛋胺酸是第一限制,離胺酸是第二限制。而當日糧蛋白質濃度低到11%和12%時,異白胺酸、蘇胺酸和色胺酸則成為限制性。
表-5:在蛋白偏低的以玉米和大豆為主要原料的飼料中,限制性氨基酸的排列順序
| 肉雞(0-3周) | 火雞(0-4周) | 生豬(10-40周) | |
| 蛋白水平(%) | 16 | 22 | 11 |
| 第一限制 第二限制 第三限制 第四限制 |
蛋氨酸 賴氨酸 精氨酸 蘇氨酸、纈氨酸 |
蛋氨酸 賴氨酸
|
賴氨酸 色氨酸 蘇氨酸 纈氨酸 |
註:在蛋氨酸和賴氨酸之後,其它限制性氨酸的排列順序暫且不明。 |
|||
離胺酸是生長豬所需的第一限制性胺基酸,色胺酸緊排在離胺酸之後,蘇胺酸是第三限制性,而且其限制程度幾乎等同於色胺酸。纈胺酸在日糧蛋白中含量低於11%,也可能成為限制性胺基酸。
在限制性胺基酸方面所做的工作很明顯地表明:可觀數量的飼料蛋白質能被幾種缺乏限制性胺基酸所取代。然而,最佳生長效果往往不可能完全被添加幾種或全部限制性胺基酸。和添加非蛋白氮的方法取得。這種例子包括將0-3週的肉雞日糧蛋白濃度由24%減到16%,將4-7週育肥階段肉雞日糧蛋白由20%減到16%,這些減少部分的蛋白質用添加各種限制性胺基酸和非蛋白氮的方法是達到不到最佳生長效果的,這其中的奧妙是目前研究的焦點。
5. 原料中胺基酸的組成
常用原料中胺基酸的組成情況列在表-6,原料蛋白質水平不能很好地反應其胺基酸組成的生物效價和被動物利用的情況。各種原料中胺基酸的平衡情況即使在蛋白質濃度極其相近的條件下也很不一樣。舉例說明:秘魯魚粉(蛋白:60%)的胺基酸平衡非常好,而玉米麵筋(蛋白:60%)的胺基酸平衡就很不理想,因為缺乏離胺酸和色胺酸。所以,分析原料中胺基酸的組成對於了解原料蛋白質質量極為重要。隨著胺基酸分析技術水準的提高,營養學家們能很容易且很快地掌握各種原料胺基酸組成的信息。
表-6 常用原料中胺基酸的百分含量
| 原料 | 蛋白 | 精氨酸 | 賴氨酸 | 蛋氨酸 | 胱氨酸 | 色氨酸 | 異亮氨酸 | 蘇氨酸 | 纈氨酸 |
| 苜蓿草葉粉 | 20 | 0.90 | 0.90 | 0.32 | 0.25 | 0.33 | 0.85 | 0.76 | 1.00 |
| 苜蓿粉 | 17 | 0.71 | 0.75 | 0.26 | 0.21 | 0.28 | 0.66 | 0.72 | 0.85 |
| 大麥(中西部) | 11.5 | 0.59 | 0.39 | 0.18 | 0.22 | 0.15 | 0.40 | 0.40 | 0.57 |
| 大麥(太平洋岸) | 9.5 | 0.44 | 0.27 | 0.14 | 0.20 | 0.12 | 0.40 | 0.33 | 0.46 |
| 血粉 | 85 | 3.60 | 6.90 | 0.95 | 0.90 | 1.15 | 0.90 | 3.90 | 7.50 |
| 干啤酒糟 | 27 | 1.30 | 0.88 | 0.45 | 0.38 | 0.37 | 1.44 | 1.00 | 1.70 |
| 啤酒酵母 | 45 | 2.20 | 3.25 | 0.70 | 0.50 | 0.50 | 2.10 | 2.10 | 2.30 |
| 菜籽餅粉 | 38 | 2.32 | 2.27 | 0.68 | 0.47 | 0.44 | 1.45 | 1.71 | 1.83 |
| 玉米 | 8.6 | 0.41 | 0.24 | 0.19 | 0.20 | 0.07 | 0.30 | 0.31 | 0.40 |
| 玉米(高賴氨酸) | 10.1 | 0.67 | 0.40 | 0.13 | 0.16 | 0.11 | 0.36 | 0.40 | 0.57 |
| 玉米胚餅粉 | 20 | 1.30 | 0.90 | 0.38 | 0.42 | 0.18 | 0.70 | 0.90 | 1.10 |
| 玉米麵筋飼料 | 22 | 1.12 | 0.72 | 0.40 | 0.51 | 0.13 | 0.70 | 0.83 | 1.15 |
| 玉米麵筋粉 | 60 | 2.00 | 1.00 | 1.70 | 1.15 | 0.35 | 2.60 | 2.00 | 2.90 |
| 玉米發酵後浸出物 | 23 | 1.00 | 0.78 | 0.50 | 0.70 | 0.05 | 0.70 | 0.90 | 1.20 |
| 棉籽餅粉(溶劑浸出) | 41 | 4.55 | 1.70 | 0.58 | 0.77 | 0.47 | 1.30 | 1.30 | 1.76 |
| 燒酒糟殘液(穀物類) | 27 | 1.00 | 0.72 | 0.55 | 0.30 | 0.20 | 1.40 | 1.00 | 1.50 |
| 燒酒糟殘液乾燥物 | 28 | 1.00 | 0.87 | 0.57 | 0.40 | 0.20 | 1.25 | 1.00 | 1.50 |
| 面包屑 | 9.5 | 0.40 | 0.26 | 0.17 | 0.20 | 0.12 | 0.45 | 0.30 | 5.80 |
| 羽毛粉 | 85 | 5.42 | 1.67 | 0.42 | 4.00 | 0.50 | 3.26 | 3.43 | 5.57 |
| 魚粉(Anchovy) | 63.5 | 3.80 | 5.00 | 1.95 | 0.65 | 0.78 | 3.00 | 2.50 | 3.23 |
| 魚粉(Menhaden) | 62 | 3.79 | 4.83 | 1.78 | 0.56 | 0.68 | 2.40 | 2.50 | 3.00 |
| 魚粉(金槍魚) | 52 | 3.30 | 4.10 | 1.36 | 0.54 | 0.62 | 2.20 | 2.30 | 2.70 |
| 濃縮魚膏 | 31 | 1.60 | 1.70 | 0.50 | 0.30 | 0.30 | 1.00 | 0.86 | 1.10 |
| 玉米麩 | 10 | 0.50 | 0.36 | 0.22 | 0.24 | 0.10 | 0.40 | 0.38 | 0.50 |
| 肉骨粉 | 50 | 3.50 | 2.65 | 0.70 | 0.63 | 0.30 | 1.50 | 1.75 | 2.35 |
| 高粱 | 9 | 0.37 | 0.22 | 0.16 | 0.18 | 0.08 | 0.36 | 0.31 | 0.48 |
| 燕麥(中西) | 11.5 | 0.77 | 0.39 | 0.18 | 0.35 | 0.13 | 0.47 | 0.40 | 0.60 |
| 燕麥(太平洋岸) | 9 | 0.60 | 0.33 | 0.13 | 0.17 | 0.12 | 0.37 | 0.28 | 0.50 |
| 去殼燕麥 | 16 | 0.90 | 0.50 | 0.20 | 0.20 | 0.18 | 0.55 | 0.43 | 0.75 |
| 花生粉(帶殼) | 42 | 5.00 | 1.60 | 0.47 | 0.70 | 0.52 | 1.70 | 1.25 | 1.80 |
| 家禽副產物粉 | 58 | 4.00 | 2.70 | 1.15 | 0.69 | 0.53 | 2.10 | 2.00 | 2.75 |
| 米糠 | 12.5 | 0.90 | 0.55 | 0.28 | 0.25 | 0.12 | 0.45 | 0.47 | 0.75 |
| 細米糠 | 12.0 | 0.50 | 0.52 | 0.20 | 0.13 | 0.10 | 0.40 | 0.40 | 0.70 |
| 粗米(Rice Rough) | 7.3 | 0.62 | 0.25 | 0.15 | 0.10 | 0.10 | 0.35 | 0.30 | 0.50 |
| 紅花籽餅粉 | 22 | 1.80 | 0.68 | 0.33 | 0.35 | 0.23 | 0.69 | 0.65 | 1.00 |
| 紅花籽餅粉 | 42 | 3.50 | 1.30 | 0.69 | 0.67 | 0.60 | 1.50 | 1.40 | 2.00 |
| 大豆粉 | 44 | 3.30 | 2.80 | 0.65 | 0.69 | 0.63 | 1.90 | 1.70 | 1.90 |
| 去殼豆粉 | 48 | 3.60 | 3.16 | 0.72 | 0.73 | 0.64 | 2.10 | 1.90 | 2.20 |
| 大豆 | 38 | 2.80 | 2.35 | 0.52 | 0.60 | 0.49 | 1.90 | 1.52 | 1.80 |
| 葵籽餅粉(帶殼) | 28 | 2.30 | 1.10 | 0.65 | 0.50 | 0.35 | 1.20 | 1.00 | 1.40 |
| 葵籽餅粉(帶殼) | 34 | 2.85 | 1.32 | 0.72 | 0.55 | 0.35 | 1.35 | 1.35 | 1.90 |
| 脫脂奶粉 | 33 | 1.20 | 2.80 | 0.84 | 0.33 | 0.43 | 1.35 | 1.55 | 2.30 |
| 小黑麥 | 15 | 0.90 | 0.50 | 0.20 | 0.30 | 0.15 | 0.60 | 0.57 | 0.80 |
| 小麥(硬) | 12.5 | 0.54 | 0.36 | 0.20 | 0.30 | 0.15 | 0.50 | 0.38 | 0.50 |
| 小麥(軟) | 10 | 0.40 | 0.30 | 0.15 | 0.22 | 0.12 | 0.45 | 0.32 | 0.44 |
| 小麥麩 | 14.5 | 1.00 | 0.58 | 0.17 | 0.35 | 0.29 | 0.50 | 0.50 | 0.70 |
| 小麥胚粉 | 24 | 1.80 | 1.40 | 0.42 | 0.46 | 0.30 | 0.90 | 0.97 | 1.10 |
| 粗小麥 | 15.5 | 1.15 | 0.70 | 0.26 | 0.37 | 0.20 | 0.60 | 0.54 | 0.70 |
| 全乳清粉 | 12 | 0.34 | 0.94 | 0.19 | 0.33 | 0.17 | 0.67 | 0.71 | 0.61 |
通過表-7可以知道為甚麼離、色、蘇和蛋胺酸在肉豬、家禽飼料中是最限制性的胺基酸。針對動物胺基酸需求量,大多數原料均缺乏這些胺基酸。表-6所列的數據是從不同資料來源綜合計算出的平均值。必須指出的是蛋白質和胺基酸的濃度即使是同一原料的不同樣品,其變化程度也相當可觀。這對某些原料來說,其意義比其它原料更重要。穀物原料如玉米、小麥和豆類原料如大豆所取不同樣本的胺基酸組成情況相對地比較穩定。可以預料,當玉米和大豆蛋白質含量增加時,其胺基酸濃度也相應地提高。然而,某些限制性胺基酸(如離、蛋胺酸等)的濃度卻不隨蛋白質的含量增加而提高。通常必需胺基酸的濃度比原料中蛋白質含量增加速度要緩慢,比較而言,某些非必需胺基酸(如:穀胺酸、絲胺酸和脯胺酸)的濃度比原料中蛋白質含量增加更快。因此,某些原料中的蛋白質往往隨著其濃度的增加而下降。
穀物和豆類原料在胺基酸組成與動物蛋白原料則相反,後者不同樣本的胺基酸組成變化很大。這些變化的主要原因一方面是使用原料方面的差異,另一方面則是加工處理方法不同。本文作者的最近研究結果證明:離胺酸在14種商業性同類肉粉中的含量變化度為2.2%-3%;胱胺酸為0.3%-1.4%;蛋白質為52%-56%,所以,胺基酸的分析對於掌握動物蛋白質質量極其重要。
6. 飼料原料中離胺酸與其他胺基酸的生物效價
前面討論過,掌握飼料原料中胺基酸的組成對日糧配方十分重要。然而,動物從飼料所能真正利用的有生物效價的胺基酸則遠遠低於飼料中所含胺基酸的量。"生物效價"一詞的含義包括消化、吸收和利用過程,即指胺基酸經吸收後能被機體利用的那部分胺基酸。掌握胺基酸的生物效價對於有效地制定出飼料配方來滿足動物的需要是很重要和必須的。
6.1 影響胺基酸生物效價的因素:
有兩種主要因素,即營養因子和各種加工處理條件。前者包括大豆中的胰蛋白𠕇抑制劑和高粱中的丹寧酸。盡量減少或排除這些抗營養因子的影響的一般方法是通過植物育種和原料的熱處理。合適的處理過程能提高原料胺基酸生物效價,然而不良的處理條件如過熱和過高壓力均能導致生物效價的降低。這些問題通常出現在那些需要加熱處理或蒸煮處理的動物性蛋白質原料和其它副產品原料。不良處理過程最容易造成離胺酸和胱胺酸效價顯著降低。前面提到過離胺酸第五位原子上的胺基團極易與還原性碳水化合物結合而形成"米勒"產物,胱胺酸在經蒸煮處理和鹼處理後能轉化成羊毛硫胺酸和其它化學物質,胱胺酸的化學轉化在羽毛粉的處理過程中常見。其它較次要的影響胺基酸生物效價的因素包括蛋白質的物理、化學性質和飼料中纖維素的含量。
6.2 胺基酸生物效價的測定:
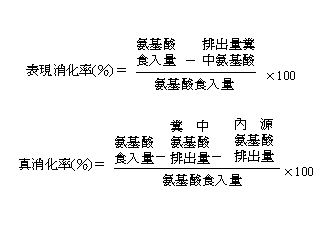
測定胺基酸生物效價的兩種主要方法是:生長試驗和消化率測定法。生長試驗法是測定某種原料取代某種特定限制性胺基酸的能力。這種原料的某種胺基酸的生物效價是通過比較這種原料和某種晶體胺基酸的生長曲線計算出來的。這種方法所需費用昂貴,人工消耗大而且通常不是很準確,每次試驗只能測定一種胺基酸。最近幾年營養學家們的興趣轉移到用消化率和平衡試驗的方法來測定胺基酸的生物效價。消化率測定的方法比生長試驗法更加簡單,而且省時、省力、省錢,特別是在每次試驗中所有胺基酸生物效價均能得到評定。消化率方法是測定食入的胺基酸和糞中(豬)或糞尿(禽)中排出的胺基酸在數量上的不同。消化率的計算方法可以是"表觀消化率"或"真消化率"。這兩者的區別在於真消化率修正了內源性胺基酸的損失。這種修正在胺基酸攝入量低時比較大,因為在這種情況下,內源性胺基酸的損失相對攝入量來說較大,表觀消化率和真消化率的計算程序列在下表。
消化率測定法的主要問題是腸道內微生物對排出胺基酸的影響。特別是大腸內微生物的降解和轉化。由於胃中和小腸內未被消化和吸收的蛋白質和胺基酸能造成胺基酸排出的數量太少,因而計算出的消化率數值偏高。為了克服這一影響因素,許多實驗室採用在豬回腸末端裝屢管或將禽的盲腸去掉的方法來測定消化率。在豬的回腸末端屢管收集消化物,能克服大腸微生物的影響,去掉盲腸後的家禽則完全消除了由此而產生的微生物的影響因素。
6.3 胺基酸消化率數據的總結
在豬禽中某些限制性胺基酸消化率系數列於表-7和表-8,表-7中家禽的消化率數據是用公禽精確飼餵法測得的。這種方法首見將成年公禽絕食24-48小時,然後將軟管插入嗉囊,通過後者餵入30-50克試驗食物。連續收集48小時的所有排瀉物(糞、尿),乾燥後,用於胺基酸的化學分析。由於所給的食物量很少,計算出的消化率被認為是真消化率。試驗家禽絕食後收集的排瀉物則用於計算內源性胺基酸的排出量。因為大多數豬的消化率都是以表觀消化率表示。所以,表-8所列值均為表觀消化率。絕大多數營養學家們都假定內源性胺基酸的損失對回腸消化率測定的影響相當小。
一般說來,家禽的消化率比豬的消化率要高,這種差別的可能原因包括:(1)在研究過程中所用原料的樣本不同:(2)兩種動物的消化率試驗方法不盡相同;(3)禽的消化是用真消化率的形式,因此可能比表觀消化率要高,如果內源性胺基酸的損失所造成的影響達到比較顯著的程度,對低於蛋白的原料(和穀物類)這後種因素大概能解釋兩種動物之間消化率的差別。
對於絕大部分原料來說,表-7所列值(家禽)均為許多樣本的平均值。這些消化率大部分是從去盲腸的家禽試驗中測得。表-8中的數據(豬)同樣也是原料樣本的平均值。來自同一原料的不同樣本,測定的消化率不一樣,浮動範圍超過20%或更高。這種現象在動物性蛋白質飼料和其它副產品的測定出現的頻率特別高。
在比較不同的飼料時,穀物類、油菜籽、玉米麵筋和血粉的胺基酸的真消化率(家禽)往往在80%以上,而其它類型的原料胺基酸的真消化率似乎比黃豆粉低10-20%。
當比較胺基酸消化率時(家禽),胱胺酸的消化率幾乎是最低的,而且比其它胺基酸低很多。許多原料中離胺酸和蘇胺酸的消化率相對也較低,蛋胺酸卻較高。
6.4 胺基酸消化率的意義和實際應用。
從表-7和-8中可以看出,所有原料中胺基酸的消化率都低於100%,而且它們的浮動範圍較大。因此,原料胺基酸濃度不能準確反應其在飼料中的相對營養價值。例如:盡管家禽副產品和去殼的大豆粉所含的離胺酸濃度一樣,但前者的離胺酸的相對效價低於後者,因其消化率本身就低。然而,晶狀胺基酸的相對營養值比原料中相對應的胺基酸要高,前者的消化率可高達100%。
表-7 飼料原料中主要必需胺基酸的真消化率(禽)1
| 原料 | 賴氨酸 | 蛋氨酸 | 胱氨酸 | 蘇氨酸 | 精氨酸 | 異亮氨酸 | 纈氨酸 |
| 苜蓿 | 62 | 72 | 36 | 67 | 80 | 78 | 76 |
| 大麥 | 76 | 84 | 80 | 78 | 85 | 82 | 83 |
| 血粉 | 85 | 92 | 76 | 88 | 87 | 79 | 88 |
| 玉米 | 79 | 90 | 82 | 84 | 89 | 87 | 87 |
| 玉米麵筋飼料 | 71 | 83 | 64 | 74 | 86 | 80 | 81 |
| 玉米麵筋粉 | 92 | 97 | 92 | 93 | 94 | 96 | 93 |
| 羽毛粉 | 67 | 75 | 57 | 74 | 85 | 87 | 85 |
| 魚粉 | 88 | 91 | 81 | 88 | 93 | 92 | 93 |
| 肉粉 | 78 | 84 | 58 | 77 | 83 | 82 | 83 |
| 燕麥 | 82 | 85 | 82 | 82 | 90 | 86 | 87 |
| 家禽副產品 | 80 | 80 | 52 | 78 | 85 | 81 | 81 |
| 油菜籽 | 76 | 89 | 70 | 76 | 88 | 83 | 80 |
| 米糖 | 70 | 74 | 60 | 68 | 83 | 73 | 72 |
| 大豆粉 | 90 | 91 | 83 | 89 | 92 | 91 | 89 |
| 葵籽粉 | 83 | 89 | 81 | 84 | 93 | 84 | 83 |
| 小麥 | 80 | 88 | 85 | 81 | 87 | 86 | 83 |
| 麥糠 | 74 | 84 | 82 | 76 | 88 | 82 | 79 |
註:以上數值是根據快速48小時排泄物收集法測定。資料主要來源於作者實驗室結果和SIBBACO(1986)和GREEN(1987)文獻。 |
|||||||
表-8 飼料原料中主要必需胺基酸的小腸末端表觀消化率(豬)1
| 原料 | 賴氨酸 | 色氨酸 | 蘇氨酸 | 蛋氨酸 |
| 大麥 | 70 | 73 | 68 | 80 |
| 血粉 | 81 | ─ | 82 | ─ |
| 油菜籽 | 72 | ─ | 67 | 84 |
| 玉米 | 70 | 65 | 65 | 84 |
| 玉米麵筋飼料 | 80 | 85 | 64 | 80 |
| 棉花籽餅粉(溶劑浸出) | 62 | 70 | 68 | 73 |
| 羽毛粉 | 50 | ─ | 74 | 64 |
| 血粉 | 85 | ─ | 84 | 87 |
| 肉粉 | 70 | 50 | 65 | 75 |
| 去殼燕麥 | 82 | 81 | 78 | 89 |
| 燕麥 | 58 | 59 | 53 | 75 |
| 花生粉 | 79 | ─ | ─ | ─ |
| 家禽副產品 | 75 | 51 | 68 | 79 |
| 黑麥 | 66 | 62 | 60 | 78 |
| 高粱 | 80 | 75 | 73 | 85 |
| 大豆粉(去殼) | 85 | 77 | 76 | 87 |
| 大豆粉(溶劑浸出) | 87 | 81 | 77 | 86 |
| 小黑麥 | 82 | ─ | 74 | 85 |
| 葵籽粉 | 74 | ─ | 72 | 86 |
| 小麥 | 75 | 78 | 71 | 85 |
| 小麥麩 | 55 | 60 | 50 | 73 |
根據胺基酸消化率制定的飼料配方應該在生產效率和經濟效益方面更好地滿足動物生長的需要,飼料的配方具有更加穩定的營養價值。幾乎所有的胺基酸需要都是以飼料中含有濃度表示,而且大多數胺基酸的消化率都是從玉米和大豆為主要原料的飼料中測定的。當使用其它原料時,飼料中胺基酸的濃度應做相應修改,因為不同原料中胺基酸的消化率不同。
許多研究結果證明:即使用蛋白質較低的飼料,只要配方是根據胺基酸消化率制定,最佳生長速度同樣能達到。
離胺酸和其它胺基酸在動物營養中應用的前景非常廣闊。人們期待利用先進的生物工程技術進一步提高離胺酸的生產效率。因此,飼料級離胺酸的產量會更高,價格更價宜。其它飼料級胺基酸也將以極具競爭性的價格進入市場。胺基酸分析技術也會不斷得到發展,將會有更多的實驗室為飼料工業界提供這項服務,隨著胺基酸各方面的資料的不斷積累,更新的數學模式將會用於預測原料中胺基酸的含量和消化率。我們應該更進一步了解低蛋白飼料中胺基酸限制的順序情況,以便通過添加這些胺基酸來取得更多的胺基酸消化率方面的資料。
所有這些都將在提高胺基酸在豬禽的利用方面得到積極的推動作用。
飼料營養雜誌(p.49∼68)─全能營養技術公司、九八年第三期